Abstract
-
Objective
- Growing hypotheses indicate the galvanic vestibular stimulation (GVS) as an alternative method to manage the symptoms of parkinson’s disease (PD). GVS is easy and safe for use, and non-invasive. However, it is elusive how the neural information caused by GVS is transmitted in the central nervous system and relieves PD symptoms. To answer this question, we investigated the transmission of neural information by GVS in the central vestibular system, focused on vestibular nucleus (VN).
-
Methods
- Twenty guinea pigs were used for this study for the extracellular neuronal recordings in the VN. The neuronal responses to rotation and GVS were analyzed by curve-fitting, and the numerical responding features, amplitudes and baselines, were computed. The effects of stimuli were examined by comparing these features.
-
Results
- Twenty six vestibular neurons (15 regular and 11 irregular neurons) were recorded. Comparing the difference of baselines, we found the neural information was linearly transmitted with a reduced sensitivity (0.75). The linearity in the neural transmission was stronger in the neuronal groups with regular (correlation coefficient [Cor. Coef.]=0.91) and low sensitive units (Cor. Coef.=0.93), compared with those with irregular (Cor. Coef.=0.86) and high-sensitive neurons (Cor. Coef.=0.77).
-
Conclusion
- The neural information by GVS was linearly transmitted no matter what the neuronal characteristics were.
-
Keywords: Synaptic transmission; Galvanic vestibular stimulation; Vestibular nuclei
-
중심단어: 신경정보의 전달, 갈바닉 자극, 전정관련 중추신경계, 파킨슨병 증상 완화
서 론
현대인들은 고령화와 생활환경의 변화로 인해 다양한 신경관련 질환에 노출되어 있다. 현재까지 알려진 신경관련 질환은 600가지가 넘으며, 이러한 신경질환은 뇌를 비롯한 신경관련 내부기관에 비정상적인 구조적, 생화학적 혹은 전기적인 문제를 일으킴으로써 장기적인 치료를 요하게 된다. 일반적으로는 약물을 이용한 치료가 주를 이루지만, 의공학적인 접근들 또한 주요한 치료 방법으로 이용되고 있다. 일례로, 대표적인 신경퇴행성 질환인 파킨슨 질환의 경우, 질환의 초기단계에는 약물에 의한 치료가 진행되지만, 질환의 말기에 이르러서는 뇌심부자극술(deep brain stimulation)에 의한 시술이 적용된다[1-3]. 이러한 전기자극을 이용한 치료법은 비정상적인 운동조절 능력과 같은 외부적인 증상에 대한 개선이라는 점에서는 매우 효과적이지만, 수술적 치료가 필요하다는 문제점이 따른다. 최근 이에 대한 대안으로 갈바닉 자극(galvanic vestibular stimulation, GVS)을 이용한 파킨슨 질환관련 증상완화에 대한 연구 보고가 점차 증가하고 있다[4-7]. GVS는 전류를 이용한 자극 방법으로 기본적인 적용방법은 한 세기 이전에 제안되었다[8]. 하지만, 자극부위에 대한 정확성이나 자극신호의 중추신경계 내에서의 전달방식은 아직도 논란을 일으킨다[9,10].
일반적으로 GVS는 전극의 연결과 전류의 극성에 따라 세 가지의 자극방식으로 나뉘게 된다. 양쪽 귀의 뒷편에 극성이 다른 전극을 연결해서 자극하는 양측이극(bilateral bipolar) 방식, 양쪽 귀의 뒷편에 동일한 극성을 갖는 전극을 연결하여 자극하는 양측단극(bilateral unipolar) 방식, 그리고 한쪽 귀의 뒷편에 자극전극을 연결하고 이마 혹은 척추쪽에 기준전극을 연결하는 단측(unilateral) 방식이 대표적이다[8,11]. 현재까지 알려진 GVS의 신경 전달적 측면에서의 영향을 보면, GVS는 반고리관내에 있는 내림프액에는 영향을 미치지 않고, 제8뇌신경인 전정-와우 신경에 직접적인 영향을 준다[12]. 또한, 불규칙적인 발화를 하는 전정관련 신경들의 발화율 변화가 규칙적인 발화를 하는 신경들에 비해 크다는 점[13]과 GVS에 의한 영향은 전정관련 신경들의 활성으로 나타난다는 점[14] 등은 GVS와 관련된 신경과학적인 발견들이다.
하지만, 이러한 단편적인 GVS에 의한 신경반응은 치료 목적으로 사용하기에는 한계성을 가진다. 이에 본 연구에서는 회전 운동자극과 GVS를 이용한 전기생리학적 동물 실험을 통해 신경정보의 전달에 대한 GVS의 영향을 알아보았다. 전기자극에 의한 영향력이 말초신경계에서는 선형적으로 이루어진다는 사실은 잘 알려져 있지만, 전정관련 중추신경계에서도 이러한 선형적인 신경정보의 전달이 이루어지는지에 대해서는 아직 실험적으로 밝혀진 바가 없다. 유사한 움직임 주파수를 갖는 회전 운동자극과 일정한 크기를 갖는 GVS를 이용하여 전정기관의 중추에 해당하는 전정핵(vestibular nucleus)에서의 전기자극에 대한 신경반응의 영향력을 평가하였다. 주어진 자극에 의해 유발된 신경정보는 곡선 맞춤(curve-fitting)을 이용하여 자극들에 의한 영향성 평가를 위한 변수들을 수치화하였으며, 이들의 비교를 통해 단위 신경에서의 반응들을 비교하였다.
대상 및 방법
20마리의 정상 기니픽(484–604 g, 수컷)를 이용하여 전정신경신호를 측정하였으며, 모든 실험 과정은 인하대학교 동물윤리위원회의 허가를 받았다.
1. 실험동물의 준비
실험동물은 케타민(0.6 mL/g)과 자일라진(0.2 mL/g)의 혼합용액을 이용하여 마취하였고, 지속적인 모니터링을 통하여 실험이 진행되는 동안 마취 상태를 유지하였다. 수술과 실험 동안에 동물의 머리 부분은 전동형의 뇌정위고정장치(stereotaxic apparatus, NEUROSTAR, Tubingen, Germany)에 의해 고정되었다. 수술은 먼저 두개골(cranium)의 상층부를 노출시키고, 노출된 부위에 양측 귀와 시상봉합선(sagittal suture line)에 의한 중심점을 이용하였다. 이 중심점을 기준으로 뒤쪽으로 1.5 mm 그리고 측면(우측)으로 2.0 mm의 위치에 측정전극의 삽입을 위한 지름 2.0 mm 크기의 입구를 열었다. 전극을 위한 입구가 완성된 후, 측정 신호의 기준(reference)을 위해 작은(지름<5 mm) 나사못을 전극입구로부터 1.5–2.0 cm 떨어진 지점에 고정시켰다. 측정전극과 관련한 준비가 끝나면 뇌경막(dura matter)을 제거하고 젖은 솜을 이용하여 열려있는 뇌 부위가 건조해 지지 않도록 덮어두었다. 모든 수술과정이 끝나면 준비된 동물은 뇌정위고정장치에 부착된 상태로 회전이 가능한 판 위에 위치시켰다.
2. 신경신호의 측정
전정핵에서의 단일 신경 신호는 에폭시(epoxy)로 코팅이 된 텅스텐 전극(5–7 MΩ, A-M Systems, Sequim, WA, USA)을 이용하여 측정되었다. 측정전극은 컴퓨터와 연결된 전동형 뇌정위고정장치의 미세한 수직적인 움직임(최소 10 ㎛)을 이용하여 목표로 하는 신경에 접근하였으며, 전극의 이동 시에 나타나는 신경신호의 특징들은 현재 전극의 위치를 알려주는 간접적인 지표로 이용하였다. 이와 함께, 전극의 수치화된 공간적 위치를 기록하여 신경신호의 측정 후에 신경의 위치를 재검토하였다. 측정 위치에 대한 재검토는 측정된 신호가 목표로 하였던 전정핵에서 측정이 되었는지에 대한 객관적인 자료를 제시함으로써 측정된 신경신호의 원천에 대한 논란이 없도록 하였다. 신경신호의 원천(수평 반고리관)은 회전자극을 이용하여 실시간으로 검사를 하였으며, 본 연구에서는 평형 회전자극에 반응하는 신경들 중에서 전기자극에도 반응하는 신경들만 사용하였다. 측정 후 신경반응의 해석을 위해, 연속된 단일 신경 발화간 시간 차이의 역수를 이용하여 순간적 발화(instantaneous firing rates, IFR)를 계산하여 자극의 유무에 따른 신경반응을 비교/분석하였다. 측정된 신경반응은 신호의 증폭(×250)과 필터링(밴드패스 필터, 0.5–3 kHz) 과정을 거쳐 저장(샘플링 레이트: 40 kHz)되었다.
3. 신경 자극 방법
본 연구에서 사용되는 주요 자극형태는 2가지로서, 평형 상태에서의 머리 회전을 이용한 운동자극과 일반적으로 GVS라 불리는 직류전류를 이용한 전기자극이다. 먼저, 자극용 전극을 측두골(temporal bone)과 측두근(temporal muscle) 사이에 위치시킨 후, 회전자극에 반응하는 신경에 양측양극 GVS를 적용하였다. 신경 발화를 활성화시키는 전기의 극성은 음극이 일반적이나, 자극용 전극의 위치에 따라 양극에 의해서 신경의 발화율이 증가하기도 하며, 이는 이전 실험에서도 증명된 바 있다[15]. 일반적으로, 동물실험에서 와우창에 GVS가 직접 적용될 때 사용되는 전류의 크기는 20–80 µA에 이르는 것으로 알려져 있다[14-16]. 하지만, 자극 위치가 와우창에서 멀어질수록 같은 수준의 신경자극을 위해서는 전류의 세기는 커져야 하며[15], 동시에 자극에 의한 통증은 최소화하는 전류의 크기를 유지해야 한다. 이러한 상황을 고려하여, 본 연구에서는 전류의 크기는 100 µA로 정하였으며, 그 이상의 전류크기에 대해서는 전류 세기에 대한 순차적인 증가(50 µA씩, 100 µA에서 500 µA까지 증가)에 대한 신경반응을 비교하여 전류크기에 의한 신경 반응 변화에 대한 통계적인 유의미성을 확인하였다. 통계적 방법은 95%의 유의성을 갖는 Wilcoxon signed rank test를 이용하였다.
단일 전정관련 신경에 대해 세 가지 유형의 자극들을 적용하여 신경반응을 측정하였다. 각각의 자극 유형은 평형 회전 운동자극만을 적용한 경우(자극 I), GVS만을 적용한 경우(자극 II), 그리고 이들 독립적인 자극을 동시에 적용한 경우(자극 III)로 구성하였다. 각각의 자극은 최소 5초동안 지속되었으며, 두 개의 자극 유형 사이에는 60초 이상의 휴지기를 두어 신경에 전달된 자극의 영향을 최소화하였다. 자극 I은 반복적인 좌우 회전운동으로, 이동 회전각은 좌우 12º 정도로 일정하게 유지하였으며, 움직임 주파수(motion frequency)는 0.7 Hz에서 1 Hz 사이의 저주파수대를 유지하였다. 자극 II는 100 µA 정도로 일정한 크기의 직류 전류를 측두골을 통하여 전달하였으며 자극에 대한 시냅스 피로(synaptic fatigue)가 일어나지 않는 범위 안에서 자극 시간을 조절하며 적용하였다. 자극 III은 이들 자극의 혼합된 형태로서 회전 자극을 지속시키는 상태에서 전기 자극을 적용하는 방식으로 구현하였다.
4. 신호해석
측정된 신경신호는 자체적으로 구현된 MATLAB (Mathworks, Cambridge, MA, USA) 프로그램을 이용하여 해석되었다. 먼저, 신경의 위치 정보를 이용하여 측정된 신경신호가 외측전정신경핵(lateral vestibular nucleus)에서 유래된 것인지를 평가하였다. 이전 논문[17]에서 제시된 외측전정신경핵의 위치를 기준으로 신경의 측정위치가 관련 영역의 범위에 포함되는지에 대한 여부를 판단함으로써 측정된 신경신호의 공간적 위치에 대한 평가를 진행하였다. 이러한 신경신호의 위치에 대한 평가는 실험 중 이루어지는 회전운동에 대한 신경반응에 대한 평가와 함께 측정된 신경신호가 목표로 한 전정핵 부위에서 측정되었다는 객관적인 정보를 제공하였다.
일반적으로 휴지기 동안의 전정관련 신경반응은 신경의 발화규칙성에 따라 규칙적 신경과 불규칙적인 신경으로 구분된다[14,18]. 단위 시간 동안의 신경발화 사이의 평균시간(m)과 표준편차(σ)에 의한 변동계수(coefficient of variation, CV)는 시간적인 변수에 의해 유동성을 가지게 되며, 이러한 유동성을 없애기 위해 정규화된 변동계수(CV*)를 이용하게 된다. 실험에서 측정된 신경신호에 대한 CV*는 공식화된 함수를 이용하여 계산하였다[18]. 본 연구에서는 계산된 CV*을 이용하여 측정된 신경들을 규칙적 발화 신경 그룹(CV*<0.1)과 불규칙적 발화 신경 그룹(CV*>0.1)으로 구분하였으며, 이외에도 신경발화를 증가시키는 전류의 극성과 전기자극에 대한 신경의 민감도를 이용한 구분들을 통해 단위 신경이 가지는 특징에 의한 신경반응에서의 변화를 고려하였다.
신경반응은 IFR 형태로 나타내었으며, (식 1)에 제시된 함수를 이용하여 비선형 곡선 접합 모델(curve-fitting model)을 구현함으로써 자극의 영향성을 수치적으로 비교하였다. 신경반응에 대한 곡선 접합 모델을 위해 사용된 함수는 (식 1)에서 보여지는 바와 같이 기본적인 사인함수를 이용해서 나타내었으며, GVS의 영향성은 사인함수의 크기(A)와 기준선(B)의 변화를 이용하여 평가하였다.
(식 1)에서 각각의 변수들(A, P, B)은 크기, 위상각, 그리고 발화율의 기준선을 나타낸다.
5. 신호해석의 근거
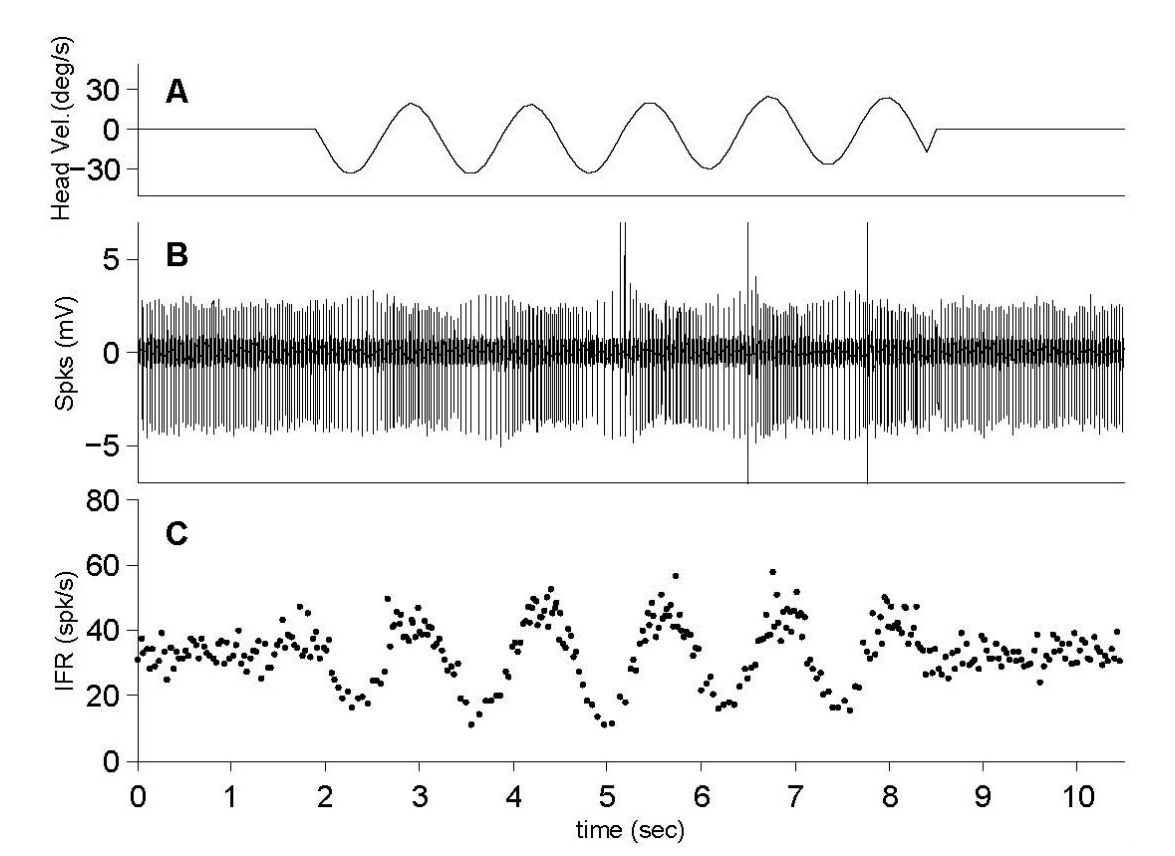
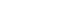
평형 회전운동과 GVS에 대한 전정관련 신경반응은 본 연구에서는 물론이고, 이전 논문들에서 자세히 언급되었다[14-16,18]. 회전운동에 대한 관련 신경들의 반응은 회전 시 발생하는 각속도의 변화에 따라 반응하게 된다(Figure 1). 각속도의 방향에 따라 신경반응이 동일한 방향성을 가지고 반응하는 신경(ipsilateral neuron)과 각속도의 방향과 반대로 반응하는 신경(contralateral neuron)이 존재하며, 이러한 각속도의 방향에 따른 신경반응은 연결된 말초기관과 관련이 있으며 자극에 대한 반응의 크기에는 영향을 주지 않는다. GVS에 의한 신경반응은 회전 운동자극에 비해 단순하다(Figure 2). 안정 전위(resting potential)는 GVS에 의해 기준선이 증가(Figure 2A) 혹은 감소(Figure 2B)된다. 이러한 기준선의 변화는 전류의 극에 따른 신경의 활성 혹은 비활성에 따른 결과로서 일반적으로는 음극의 전류에 의해 신경의 활성이 증가한다고 알려져 있다. 본 연구에서는 전기자극에 의한 신경변화에서 증가하는 신경반응만을 해석의 자료로 이용하였다.
주어진 신경 자극방법에서 자극III은 자극I과 자극II에 의한 복합적인 자극방법으로 곡선 접합 모델을 이용한 해석법에 따르면, 자극I 은 사인함수의 변수 중에 크기(A)와 위상차(P)에 영향을 주고, 자극II는 기준선에 영향을 주는 것으로 나타났다. 또한 위상차(P)는 운동자극의 회전 움직임의 빠르기에 따라 변하게 되므로, 단일자극(자극I)과 복합자극(자극III)에서 회전 움직임의 빠르기가 동일하다면 회전 운동자극에 의한 신경변화는 함수의 크기(A)로만 나타난다. 따라서, 복합적인 자극형태가 신경에 주어지더라도 자극의 영향이 함수의 변수에 중복되어 나타나지 않는다. 이러한 신경에 대한 자극의 독립적인 영향을 기초로 하여 자극II의 영향을 신경반응 함수의 크기(A)와 기준선 (B)를 이용한 비교를 통해 알아보았다. 단일 신경에서의 변화를 이용하여 신경집단에 대한 전체적인 반응추세를 나타냄으로써 신경정보 전달의 선형성에 대한 결론을 도출하고자 하였다.
자극II에 의한 기준선의 변화(변화A)와 자극I과 자극III의 기준선 변화(변화B) 사이의 선형적 관계에서의 주요성을 통계적으로 평가하기 위해 기울기의 표준오차(standard error)를 이용한 t-test를 적용하였다. 만일, 두 변화 사이에 선형적인 관계가 존재하지 않는다면 기울기는 0이 될 것이고(귀무가설, null hypothesis), 존재한다면 기울기는 0이외의 값을 가지며 선형관계를 유지할 것이다(대립가설, alternative hypothesis). 즉, t-test에 의해 p-value의 값이 귀무가설을 기각하게 되면(p-value<0.05), 변화A와 변화B사이에는 선형적인 관계가 존재한다고 할 수 있다.
결 과
20마리의 기니픽을 이용하여 26개의 전정관련 신경반응을 측정하였으며, 발화율의 규칙성을 바탕으로 한 분류[14,18]를 통해 15개의 규칙성 발화를 하는 신경들과 11개의 불규칙 발화를 하는 신경들로 구분하였다. 측정된 신경반응의 전기자극의 극성에 따른 분포에 따르면(Figure 3), 음극에 의한 발화율 증가가 우세하나(p=0.015, Binomial cumulative distribution) 양극에 의한 신경 발화율의 증가 또한 존재함을 알 수 있었다. 회전 방향에 따른 신경 구분에서, 동측 방향에서 나타나는 신경반응의 증가는 그 반대 경우에서 나타나는 신경반응의 증가에 비해 상대적으로 적었다(p=0.038, Binomial cumulative distribution). 이에 반해, 발화율의 규칙성과 자극에 대한 민감도에 따른 신경의 구분에서는 그룹 간 차이의 통계적인 유의미성은 나타나지 않았다(p>0.084, Binomial cumulative distribution). 직류전류의 크기에 따른 신경반응의 변화를 보기 위해 전류의 크기를 달리하면서(100–500 µA) 그에 따른 신경반응을 확인하였다. 전류의 크기는 100 µA에서 시작하여 50 µA씩 상승시키며 그에 따른 신경반응의 변화를 측정하였으며, 자극 시에 나타나는 신경의 IFR를 비모수적 검정을 이용하여 비교하였다. 통계적 방법을 이용한 비교에서 전류크기의 증가는 신경반응에 영향을 주지 못하였다(p>0.21, Wilcoxon signed rank test). 이는 100 µA에 의한 GVS가 그 이상의 자극 크기를 이용한 신경반응에서도 유사하게 나타남을 의미하며, 자극강도에 있어서 충분한 자극이 적용되었음을 의미한다.
모든 단일신경에 세 가지 형태의 자극이 주어졌으며, 각각의 자극에 대한 신경반응을 측정하였다. 단일 자극에 의한 영향이 신경반응에 지속적으로 영향을 주는지 알아 보기 위해 휴지기 동안의 발화율과 GVS 이전의 발화율을 비교하였다(Figure 4). 일반적으로 회전자극에 의한 시냅스피로(synaptic fatigue)는 관찰되지 않으나, 5초 이내의 짧은 시간 동안의 GVS에서는 시냅스 피로가 관찰되었다. 이전에 주어진 자극의 영향은 독립된 자극에 의한 신경반응 해석에도 영향을 주게 되므로, 단위 시간 동안의 휴지기와 GVS 전의 신경 발화율 차이를 비교함으로써 앞선 자극의 영향을 측정하였다. 다수(21/26, 80.8%)의 발화율 차이는 휴지기 동안의 발화율 범위(4–62 spikes/sec)보다 낮았으며, 이는 GVS에 의한 영향이 다음에 오는 자극 이전에 소멸되었음을 의미한다. 평균값에 의한 비교와 함께 통계적인 방법을 이용한 발화율의 차이를 검증하였으며, 휴지기 동안과 GVS 이전의 모든 신경반응에서 발화율에는 차이가 없다는 결과를 얻었다(p>0.11, Wilcoxon signed rank test).
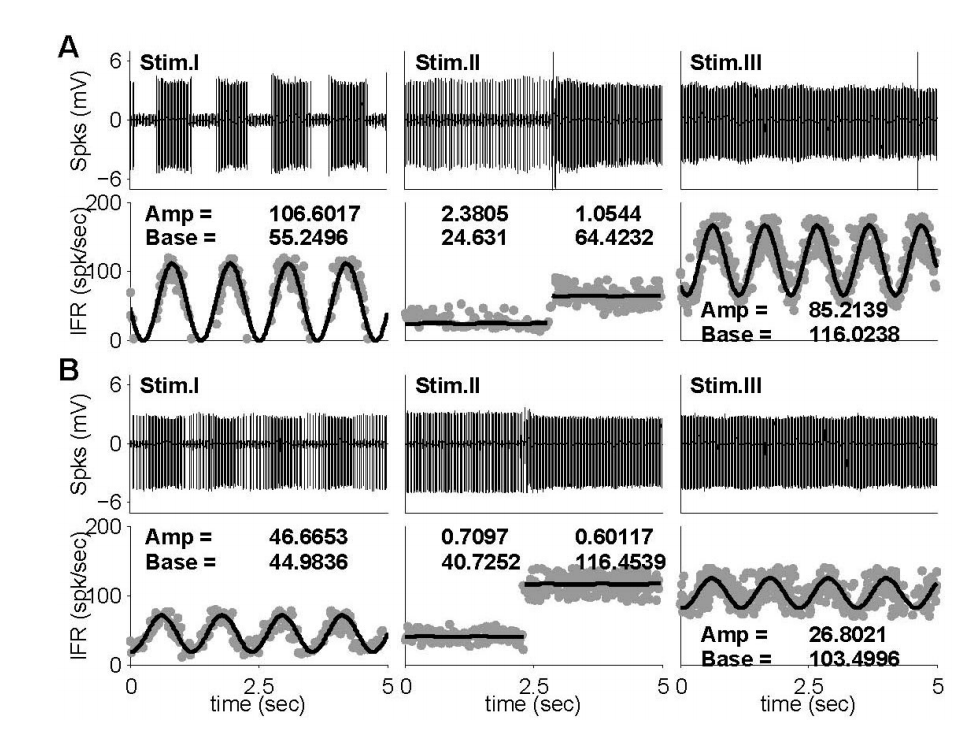
측정된 자극 대비 신경반응은 IFR형태로 표현된 후에(식 1)을 통한 비선형 곡선 접합 모델을 구현하였다. 구현된 비선형 곡선 접합 모델을 이용하여 각각의 반응에서 크기(A)와 기준선(B)을 계산하였으며, 계산된 값들은 단일신경 반응 비교에 이용되었다. 설명된 신경반응 해석방법은 Figure 5에서 보여지는 단일 신경반응의 예에서 보여지듯이, 자극I에 대한 신경반응은 모델(식1)에서 제시된 사인파형을 나타내었으며, 크기와 기준선은 각각 106.60과 55.25 spk/sec였다(Figure 5A). 이에 반해, 자극II에 대한 반응은 뚜렷한 사인파형은 나타나지 않았으며, GVS의 전(24.63 spk/sec)과 후(64.42 spk/sec)에 변화된 기준선에 의해서 자극의 영향을 확인할 수 있었다. 자극III에 의한 신경반응은 자극I에 대한 신경반응과 비교할 때, 사인파형의 기준선은 55.25 spk/sec에서 116.02 spk/sec로 증가하였고, 파형의 크기는 106.60 spk/sec에서 85.21 spk/sec로 감소하였다. 단일 자극형태가 주어질 때, GVS에 의한 신경반응의 영향은 자극의 전과 후의 기준선의 변화를 통해 평가된다. 하지만, 주어진 예(Figure 5)의 자극II에 의한 신경반응의 변화는 39.79 spk/sec였으며, 이는 자극I과 자극III의 기준선 차이(60.77 spk/sec)와 비교할 때, 기준선의 변화는 증가(52.74%)하였다. 이에 반해, Figure 5B에서 보여지는 신경반응의 예는 GVS의 전(40.73 spk/sec)과 후(116.45 spk/sec)의 기준선에서의 변화는 자극I과 자극III에서의 신경반응의 기준선의 변화(58.52 spk/sec)와 비교했을 때 감소(22.72%)하였다. 신경반응의 기준선 변화는 반대로 나타났지만, 이전 신경반응의 예(Figure 5A)에서 나타났듯이 자극I의 파형 크기(46.67 spk/sec)에 비해 자극III의 파형 크기(26.80 spk/sec)는 감소하였다.
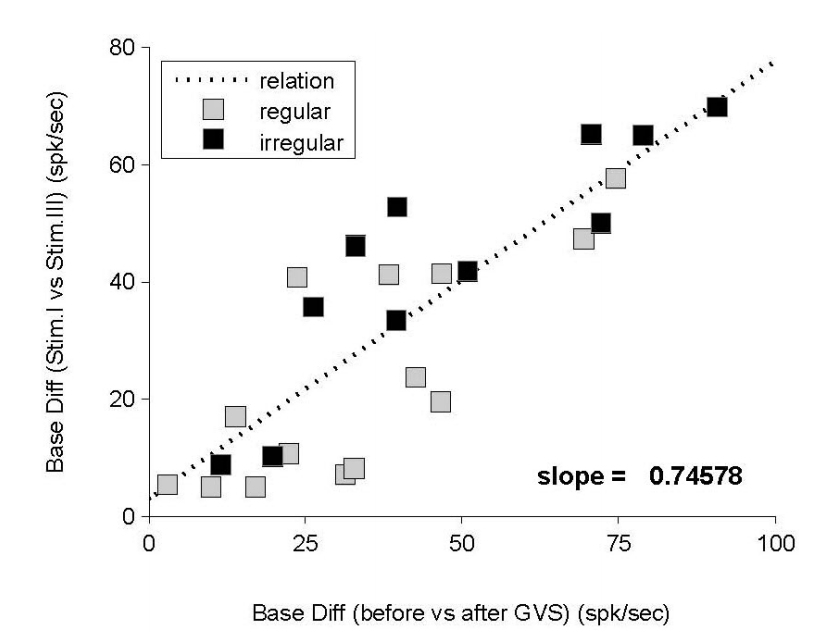
동일한 해석방법을 이용하여 측정된 모든 신경반응을 해석하였다. GVS에 의한 신경반응은 기준선의 변화로 나타나므로, 자극II에서 나타난 기준선의 변화(변화A)를 자극I과 자극III의 기준선 차이(변화B)와 비교하였다(Figure 6). 18개의 신경반응에서 변화A가 변화B보다 크게 나타났으며, 이는 대부분의 반응에서 변화A가 변화B보다 크게 나타남을 의미한다(p=0.0145, Binomial cumulative distribution). 전체적인 기준선 변화의 관계는 선형적으로 나타났으며, 통계적인 방법에 의한 평가에서도 변화A와 변화B 사이의 선형성은 유의미한 관계를 보였다(p<0.01).
고 찰
본 연구에서는 GVS에 의한 신경 반응이 전정 시스템의 중추에 해당하는 전정핵에 선형적 혹은 비선형적으로 전달되는가에 대해 계산 신경과학(computational neuroscience)적 방법을 이용하여 조사하였다. 말초 전정기관으로 입력된 GVS에 대한 신경반응은 비선형 곡선 접합 모델에 기초한 기준선의 변화를 통해 나타남을 확인하였으며, 단일자극(자극II)과 복합적인 자극(자극III)에 대한 신경반응의 기준선 비교를 통해 주어진 GVS에 의한 신경정보 전달 형태를 연구하였다. 단일자극(자극II)에서의 신경반응 기준선의 변화는 운동자극(자극I)과 복합자극(자극III) 사이에서 나타나는 기준선의 변화와 비교할 때 증가(Figure 5A) 혹은 감소(Figure 5B)하였으나, 다양한 특징들(신경 발화율, 자극 민감도, 발화 극성, 발화 방향)을 갖는 신경들을 포함하는 전체 집단에서의 기준선 변화 비교 시 GVS에 대한 신경 정보는 선형적으로 전달됨을 나타내었다. 본 연구에서 GVS의 크기는 100 µA로 한정되어 있었으나, 단일 신경의 자극에 대한 민감도는 짧은 시간 동안의 자극으로 인해 변할 수는 없으므로 전정 신경에 자극의 영향이 미치는 수준에서의 자극 크기에서는 유사한 결과가 예상된다. 또한, 자극의 영향은 기존의 논문15에서 밝힌 GVS 세기(20–60 µA & 620 µA) 대비 신경 발화율의 변화와 본 연구에서 측정한 자극 변화(100–500 µA)에 대한 신경 발화율 변화의 결과를 토대로 결정되었으며, 다른 연구[16]에서도 동일한 자극수준을 갖는 연구가 진행된 점 등을 근거로, 본 연구에서 사용된 자극의 세기 수준은 적정한 것으로 판단된다. 더불어, 자극이 100 µA에 한정되어 있으나, 단위 신경이 나타내는 민감도에서는 차이를 보였으며, 이는 신경에 대한 자극의 영향력 차이로 판단된다. 따라서, 자극에 의한 영향은 단위 신경에 다르게 작용하므로, 다수의 신경에 유사한 영향을 나타내는 단일 자극의 크기는 존재하지 않는다. 하지만, 다수의 신경들에 의해 유발되는 반응의 선형성은 대부분의 신경들이 영향을 받을 정도의 충분한 자극 크기에서 나타나는 신경반응 변화 사이의 비교를 통해 예측할 수 있으며, 본 연구에서 적용된 자극의 크기(100 µA)는 이러한 조건을 충족하는 수준으로 판단된다.
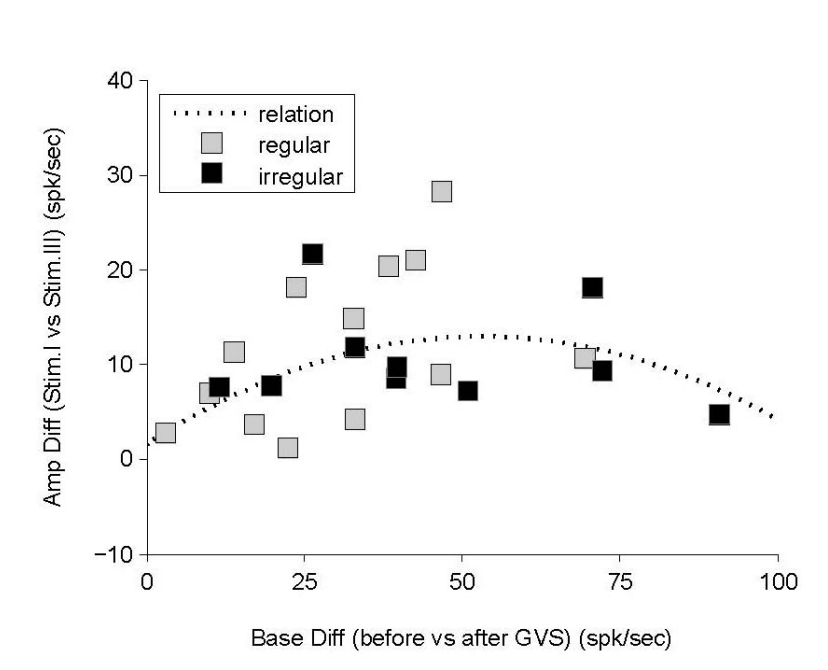
운동자극(자극I)에 의한 신경반응은 파형의 크기로 나타나게 됨을 결과(Figure 5)에서 보였다. 하지만, 운동자극에 대한 신경정보는 GVS와의 혼합(자극III)에서는 파형의 크기를 줄였으며, 이는 측정된 신경 전반(25/26, 96.2%)에서 나타나는 현상이었다. 위 결과는 GVS에 대한 신경반응 변화와 비교해 볼 때, 전정관련 신경에서는 자극의 우위가 존재함을 암시하며, GVS가 운동자극에 비해 신경반응의 영향력에 있어서 우위에 있다고 할 수 있다. 하지만, 이러한 우위성은 GVS에 대한 신경의 민감도에 따라 다르게 나타난다. GVS 동안의 신경반응의 변화에 대한 신경반응의 크기 변화에 대한 비교(Figure 7)에서 자극에 대한 민감도가 올라갈수록 복합자극(자극III)에서의 신경반응 크기는 줄어들지만, 일정 수준의 민감도(0.5 spk/sec/µA)를 넘어서면 복합자극에서의 파형 크기의 감소 또한 줄어들었다. 위 결과를 토대로 GVS의 우위성은 자극에 대한 민감도가 낮을 때 강했으며, 자극에 대한 민감도의 증가는 자극으로서의 우위성 약화를 뜻한다고 할 수 있다.
결 론
GVS는 파킨슨병 관련 증상 완화를 위한 자극 형태로서 최근 여러 연구에서 주목을 받고 있다. 이에, GVS에 의한 전정관련 신경반응을 이용하여 자극에 의한 생리학적인 영향을 알아보았다. 운동 자극과 GVS를 이용한 혼합 자극을 이용하여 각각의 독립된 자극의 신경 반응에 대한 영향을 측정 및 해석하였다. GVS의 신경반응은 기준선의 변화에 의해 나타났으며, 이에 기초하여 단일 자극과 혼합 자극에서의 신경반응의 기준선 변화를 비교하여 GVS에 의한 중추 신경계에서의 영향을 조사하였다. 신경반응의 기준선 변화를 이용한 비교를 통해, GVS에 의한 신경정보는 선형적으로 전달됨을 확인하였으며, 운동자극보다 GVS의 신경에 대한 영향이 우위에 있음이 나타났다. 다양한 크기의 자극 적용을 통하여 보다 광범위한 자극에 대한 신경정보의 선형적 전달에 대한 이해를 높일 수 있을 것으로 기대된다.
ARTICLE INFORMATION
-
No potential conflict of interest relevant to this article was reported.
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded partially by the Ministry of Education (2010-0020163) and partially by the Ministry of Education (2016R1D1A1B03930657).
Figure 1.Neural responses to horizontal head rotation in the vestibular nucleus of a guinea pig. The maximum velocity of head rotation is 30°/sec, and the neuronal response corresponds to the rotatory velocity. (A) The change of head velocity (°/sec). (B) Neuronal spikes (mV) during the head rotation. (C) Instantaneous firing rate (IFR, spk/sec) based on the spikes.
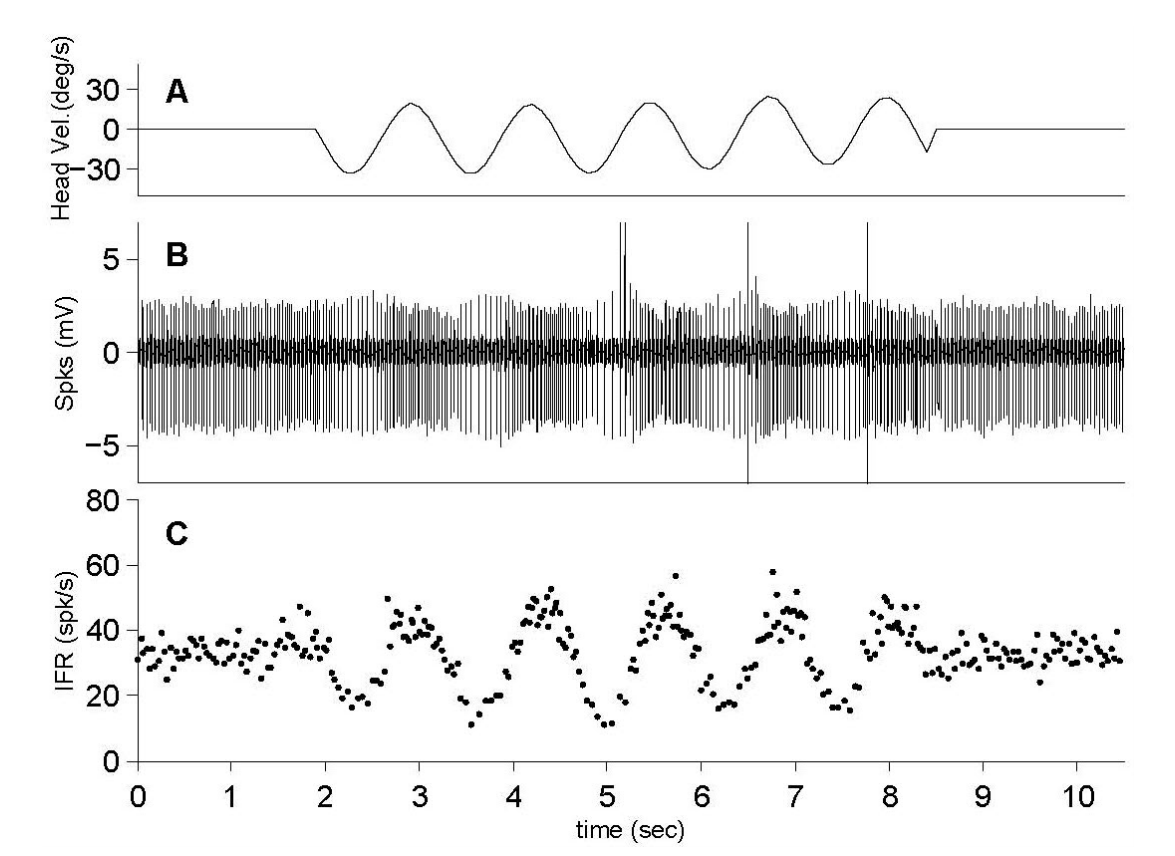
Figure 2.The exampled neuron increases its firing rate (FR) when an excitatory galvanic vestibular stimulation (GVS) is applied, and decreases its FR by an inhibitory GVS. (A) Neuronal excitation by a negative GVS. (B) Neuronal inhibition by a positive GVS from the same neuron. IFR, instantaneous firing rates.
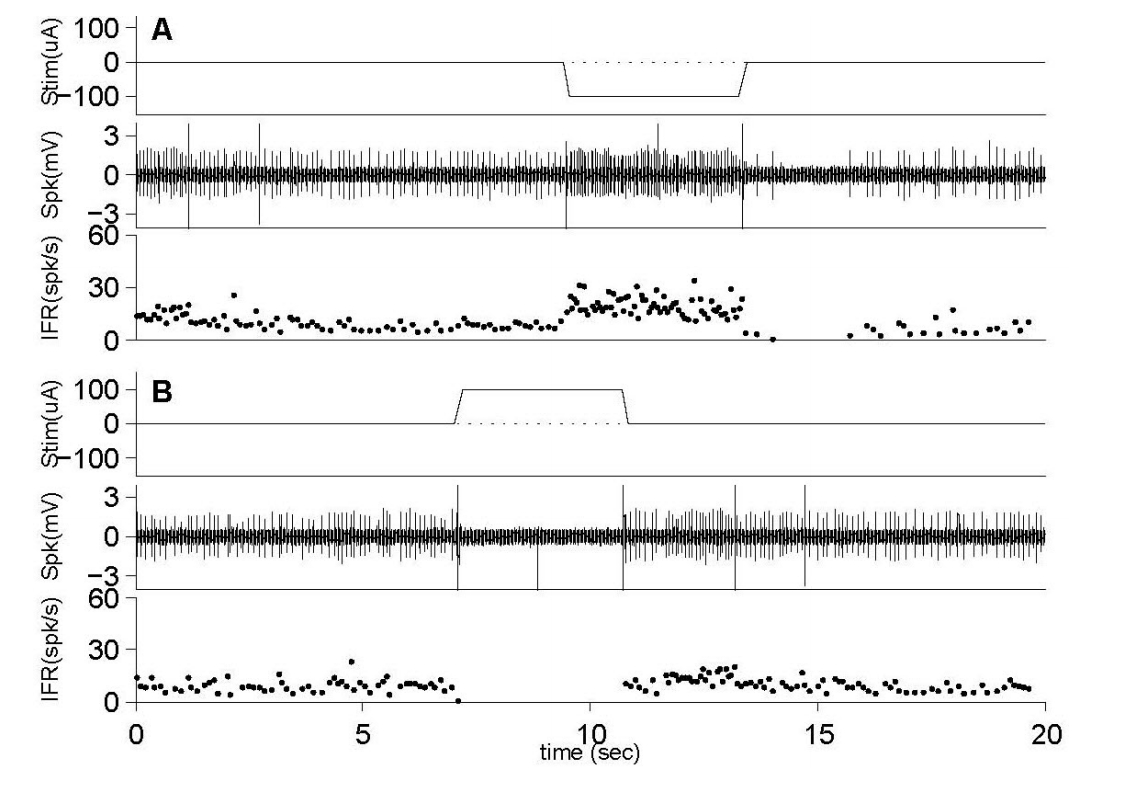
Figure 3. Classification based on the neuronal responding characteristics. (A) Classification by discharge regularity. Based on the normalized coefficient of variation (coefficient of variation, CV**), the neurons are separated regular (CV*<0.1) and irregular (CV*> 0.1) units. (B) Classification by galvanic vestibular stimulation (GVS) polarity to excite neuronal firing rate (FR). (C) Classification by neuronal sensitivity (spk/sec/µA) to GVS. The low and high sensitive groups are separated by the mean of sensitivity. (D) Classification by the rotatory direction. If neuronal FR increases as the head moves to the rightward (ipsi) direction, it is identified as ipsi-preferred neuron.
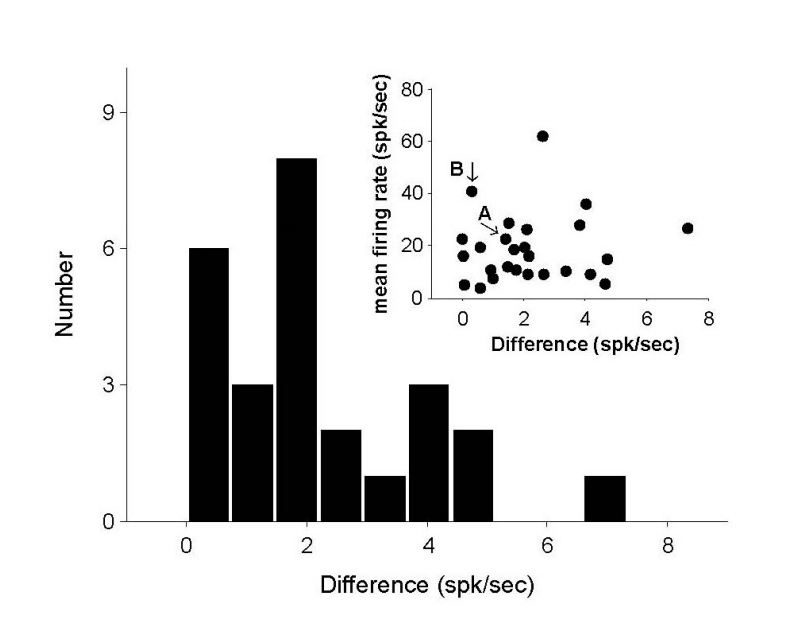
Figure 4.The difference of firing rate (FR) during no stimuli. To measure the remained effects by previous stimuli, the difference between the initial FR and the FR after several stimuli is compared. Twenty five (96.2%) neurons show lower than 5 spk/sec, and only one difference is 7.3 spk/sec. The subplot show the comparison between FR difference and mean FR. The arrows indicates the exampled units in Figure 5A and 5B.
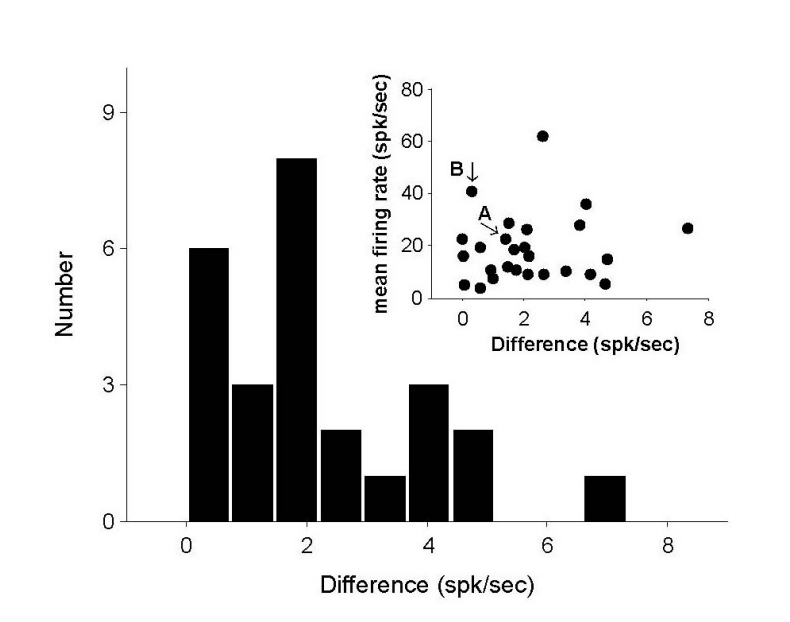
Figure 5.Two exampled neuronal responses to head rotation (Stim. I), GVS (Stim. II), and mixed stimulus (Stim. III) and their nonlinear curve-fittings. (A) The neuronal responses of an irregular (CV*=0.17), low sensitive, ipsi-preferred, and excited by a negative GVS. (B) The neuronal responses of a regular (CV*=0.04), high sensitive, contra-preferred, and excited by a positive neuron. GVS, galvanic vestibular stimulation; CV, coefficient of variation; IFR, instantaneous firing rates.
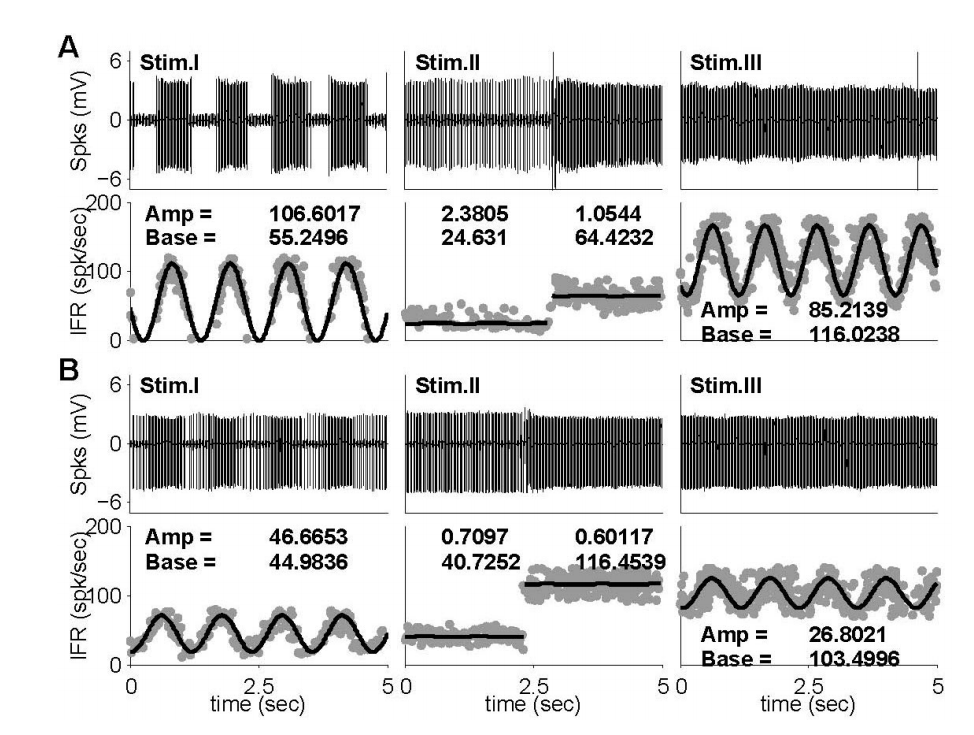
Figure 6.Relation between two baseline differences; one by GVS and the other by the baseline difference of Stim. I and Stim. III. Due to the independence between stimuli, the presented relation indicates the effect of GVS. There is a linear relation between two baseline differences, and the slope is 0.75. GVS, galvanic vestibular stimulation.

Figure 7. Relation between the baseline difference and the amplitude difference; the baseline difference is caused by GVS and the amplitude difference is the change in amplitudes during Stim. I and Stim. III. The overall relation is bell-shaped by constructing a positive and a negative linearity under the small (<50 spk/sec) and the large difference of baselines, respectively. GVS, galvanic vestibular stimulation.
REFERENCES
- 1. Krack P, Batir A, Van Blercom N, Chabardes S, Fraix V, Ardouin C, et al. Five-year follow-up of bilateral stimulation of the subthalamic nucleus in advanced Parkinson’s disease. N Engl J Med 2003;349:1925–34.ArticlePubMed
- 2. Schupbach WM, Chastan N, Welter ML, Houeto JL, Mesnage V, Bonnet AM, et al. Stimulation of the subthalamic nucleus in Parkinson’s disease: a 5 year follow up. J Neurol Neurosurg Psychiatry 2005;76:1640–4.ArticlePubMedPMC
- 3. Horn A, Kuhn AA. Lead-DBS: a toolbox for deep brain stimulation electrode localizations and visualizations. Neuroimage 2015;107:127–35.ArticlePubMed
- 4. Yamamoto Y, Struzik ZR, Soma R, Ohashi K, Kwak S. Noisy vestibular stimulation improves autonomic and motor responsiveness in central neurodegenerative disorders. Ann Neurol 2005;58:175–81.ArticlePubMed
- 5. Pan W, Soma R, Kwak S, Yamamoto Y. Improvement of motor functions by noisy vestibular stimulation in central neurodegenerative disorders. J Neurol 2008;255:1657–61.ArticlePubMed
- 6. Okada Y, Kita Y, Nakamura J, Kataoka H, Kiriyama T, Ueno S, et al. Galvanic vestibular stimulation may improve anterior bending posture in Parkinson’s disease. Neuroreport 2015;26:405–10.ArticlePubMed
- 7. Rizzo-Sierra CV, Gonzalez-Castano A, Leon-Sarmiento FE. Galvanic vestibular stimulation: a novel modulatory countermeasure for vestibular-associated movement disorders. Arq Neuropsiquiatr 2014;72:72–7.ArticlePubMed
- 8. Kim G, Lee S, Kim K-S. Vestibular system research based on electrophysiology. Korean J Otorhinolaryngol-Head Neck Surg 2016;59:83–7.Article
- 9. Cohen B, Y akushin SB, Holstein GR. What does galvanic vestibular stimulation actually activate? Front Neurol 2011;2:90. ArticlePubMed
- 10. Curthoys IS, Macdougall HG. What galvanic vestibular stimulation actually activates. Front Neurol 2012;3:117. ArticlePubMedPMC
- 11. Fitzpatrick RC, Day BL. Probing the human vestibular system with galvanic stimulation. J Appl Physiol (1985) 2004;96:2301–16.ArticlePubMed
- 12. Pfaltz CR, Koike Y. Galvanic test in central vestibular lesions. Acta Otolaryngol 1968;65:161–8.ArticlePubMed
- 13. Ezure K, Cohen MS, Wilson VJ. Response of cat semicircular canal afferents to sinusoidal polarizing currents: implications for input-output properties of second-order neurons. J Neurophysiol 1983;49:639–48.ArticlePubMed
- 14. Goldberg JM, Smith CE, Fernandez C. Relation between discharge regularity and responses to externally applied galvanic currents in vestibular nerve afferents of the squirrel monkey. J Neurophysiol 1984;51:1236–56.ArticlePubMed
- 15. Kim J, Curthoys IS. Responses of primary vestibular neurons to galvanic vestibular stimulation (GVS) in the anaesthetised guinea pig. Brain Res Bull 2004;64:265–71.ArticlePubMed
- 16. Courjon JH, Precht W, Sirkin DW. Vestibular nerve and nuclei unit responses and eye movement responses to repetitive galvanic stimulation of the labyrinth in the rat. Exp Brain Res 1987;66:41–8.ArticlePubMed
- 17. Rapisarda C, Bacchelli B. The brain of the guinea pig in stereotaxic coordinates. Arch Sci Biol (Bologna) 1977;61:1–37.PubMed
- 18. Massot C, Chacron MJ, Cullen KE. Information transmission and detection thresholds in the vestibular nuclei: single neurons vs. population encoding. J Neurophysiol 2011;105:1798–814.ArticlePubMedPMC
Citations
Citations to this article as recorded by



 PubReader
PubReader ePub Link
ePub Link Cite
Cite