Abstract
- The velocity-storage circuit comprised of bilateral vestibular nucleus complexes, commissural fiber, and nodulus and uvula functions in refining the raw vestibular signal to estimate rotational velocity, gravity direction, and inertia. In this review, we pursued the functional significance of this velocity-storage circuit and how this physiologic knowledge could help us understand the clinical symptoms and signs of patients with vestibular disorders.
-
Keywords: Cerebellum; Vestibular diseases; Dizziness; Vertigo
-
중심단어: 소뇌, 전정질환, 어지럼, 현훈
서 론
전정계는 머리의 가속도를 신경신호로 전환하는 생리적 가속도계(biologic accelerometer)이다[1]. 반고리관과 이석기관, Scarpa’s 신경절을 포함한 상ㆍ하 전정신경을 말초전정신경계라고 한다[2]. 중추전정신경계는 다소 복잡한 신경 네트워크를 형성한다. 양쪽 전정신경핵과 이를 연결하는 교차섬유(commissural fiber)를 기본으로 하며, 타래 및 부타래, 결절 및 목젖으로 대표되는 전정소뇌가 주요 구조물이다[2]. 이들 구조물은 교뇌, 중뇌, 시상 및 전정피질과는 안쪽세로다발(medial longitudinal fasciculus), 위소뇌다리(brachium conjunctivum), 배쪽뒤판로(ventral tegmental tract) 등의 상행로를 통해 연결되고, 뇌간, 척수의 자율 및 운동신경핵과는 내ㆍ외 전정척수로의 하행로를 통해 연결된다[3].
전정신경계의 기능
전정계는 말초전정신경계와 중추전정신경 네트워크를 통해 전정인식, 전정안반사, 전정(목)척수반사, 전정자율반사를 형성한다. 임상적으로 전정안반사 이외에 다른 부분에 대해 이해를 돕기 위해 세부적으로 기술하겠다.
1. 전정인식
속도(velocity)와 가속도(acceleration)에 대한 인식을 의미하며, (1) 중력에 대한 머리의 위치 인식(gravity perception in head reference frame), (2) 이동 중 발생하는 머리의 속도 인식(head velocity perception during motion), (3) 그리고 주변 환경과 연관하여 머리의 위치와 속도를 인식하는 것으로 구성할 수 있다[4]. 먼저 중력에 대한 인식이라는 것은 우리가 지구라고 하는 구체 위에 서로 다른 위치에 존재함에도 불구하고 바로 서 있다는 인식을 가능하게 해주는데, 이는 우리가 머리 방향(heading direction)보다는 지구중심을 향한 중력가속도의 반대방향을 바로 서 있는 기준으로 한다는 것을 의미한다[5]. 반면, 우주와 같이 중력이 없는 환경에서 바로 서 있다는 것의 기준은 머리 방향이 된다. 두 번째와 세 번째에 관해 설명하면, 움직이는 것은 크게 주체(나)의 움직임(subjective motion)과 객체(상대)의 움직임(objective motion)으로 구분할 수 있다. 주체의 움직임은 전정종말기관(vestibular end organ)을 통해 측정된다. 반면 객체의 절대적 움직임을 측정하는 기관은 없으며 주체와 객체의 상대적 움직임(relative motion)을 측정하는 기관은 시각과 촉각이다. 시각흐름(optic flow)은 주체와 객체의 상대적 위치를 시간에 따라 전달하므로 속도신호가 파생되며, 촉각은 압력(pressure)신호를 전달하는데, 압력(p)은 단위면적당 가해지는 힘(f)이며 힘은 다시 질량(m)과 가속도(a)를 가지고 있어 결국 시간에 따른 압력의 변화는 속도신호를 포함하고 있다. 이러한 상대적 움직임에 대한 신호는 중추신경계에서 전정신경에서 전달된 주체의 절대적 속도를 감하여(subtracting) 객체에 대한 절대적 신호를 전달하게 된다[6-8]. 따라서 머리의 움직임에 대한 정보 정확성은 주체의 올바른 속도 인식과 더불어 객체의 속도신호에 대한 정확성도 담보할 수 있다.
2. 전정(목)척수반사
전정(목)척수반사는 머리움직임 동안 머리의 위치를 공간에서 안정적으로 유지하게 한다[9]. 이는 전정안반사의 목적(눈의 위치를 공간상에서 안정적으로 유지)과 일맥상통한다. 즉 머리가 수동적(passive)으로 좌측으로 회전하면 전정척수반사는 우측으로 머리를 회전시켜 원래 위치 방향을 향하게 한다. 이를 위해 좌측 목빗근(sternocleidomastoid muscle)과 우측 척추옆근육(paraspinal muscle)을 수축시킨다. 다만 의도적 머리 회전(voluntary head motion)의 경우 전정(목)척수반사는 작동이 되지 않는다. 전정(목)척수반사와 같이 작동하는 반사가 경부-목반사(cervicocollic reflex)인데, 전정(목)척수반사와 다르게 목적은 머리 위치를 몸 위에서 안정적으로 작용하게 하는 것이다[10]. 즉 수동적으로 좌측으로 머리가 회전하면 전정(목)척수반사는 머리를 우측으로 이동시키지만, 경부-목반사는 머리를 좌측으로 이동시킨다(Fig. 1). 두 개의 반사가 쌍으로 조화롭게 작용하면서 공간과 몸에서 안정적으로 위치하게 한다[11].
3. 전정자율반사
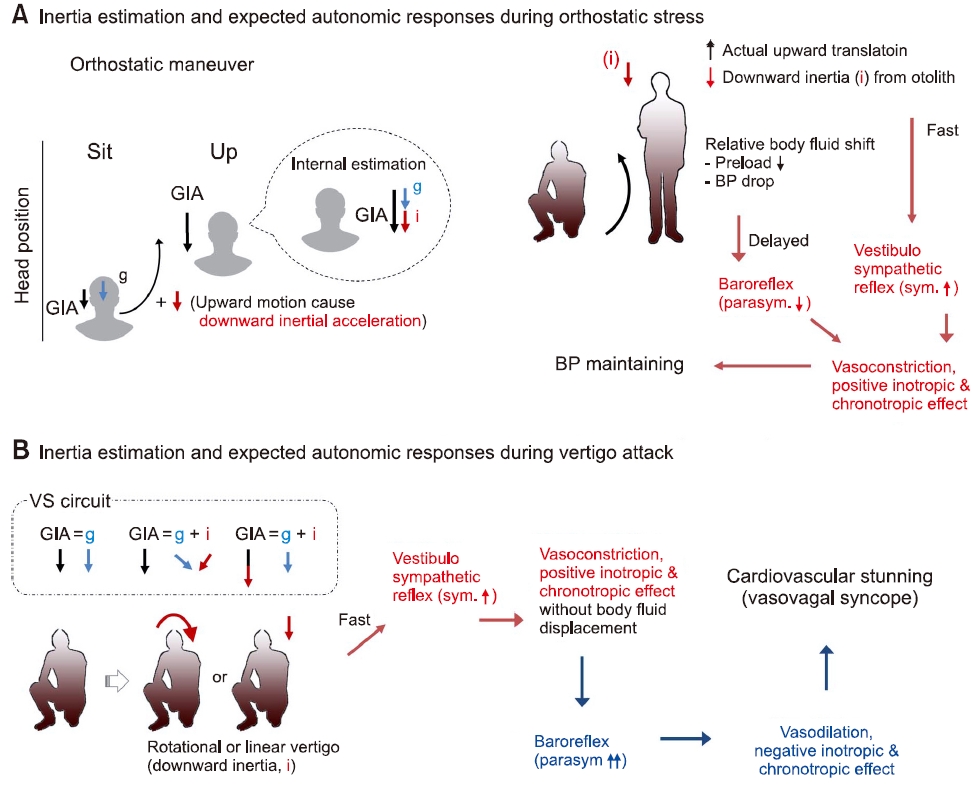
전정자율반사는 심혈관의 항상성에 관여하는 신호로서, 몸의 움직임에 동반하여 발생하는 체액의 상대적 위치 변화에 따른 혈압 변화를 보상한다. 이석기관과 교감신경계를 통해 주로 작동하므로, 전정교감반사(vestibulo-sympathetic reflex) 또는 이석교감반사(otolith-sympathetic reflex)라고 명칭되기도 한다[12,13]. 이 반사는 feedforward 형식인데, 몸의 움직임에 동반한 혈압 변동을 예측하여 교감반사의 활성을 우선 조절하며, 100–200 msec의 짧은 잠복기로 작동한다. 반면 압력반사(baroreflex)는 feedback 형식으로 작용하고 1–2초의 잠복기를 가지며 부교감 신경계의 활성에 주로 관여한다[14]. 따라서 전정자율반사와 압력반사가 상호 보완적으로 작용하여 혈압의 항상성을 유지하게 된다(Fig. 2).
말초전정신경계의 작동 원리와 불완전성
앞부분을 통해 전정계가 머리의 속도신호를 뇌에 전달하고 중추신경 네트워크를 통해 인식, 눈운동, 자세, 그리고 자율신경계에 작동하는 방식에 대해 살펴보았다. 이제 그 기능을 수행함에 있어 말초전정계의 문제점에 대해 확인해 보겠다.
1. 각속도 측정에 있어 반고리관 기능의 불완전성
반고리관은 골성미로(bony labyrinth)와 막성미로(membranous labyrinth)로 구분되며, 골성미로와 막성미로 사이에는 외림프가, 막성미로 안쪽은 내림프(endolymph)가 차지하고 있다. 머리의 움직임에 의해 막성미로와 내림프의 움직임에 상대적인 차이가 생기면 cupula는 회전반대방향으로 편위(deflection)되어 신경전위가 발생하는 원리이다. 그러나 머리가 가속도가 없이 등속도로 한 방향으로 회전하면, 내림프가 막성미로와의 마찰력에 의해 같이 회전하며 내림프와 막성미로 사이에 상대적 움직임의 차이가 점차 사라지게 된다. 따라서 cupula는 편위된 위치에서 점차 원래 위치로 돌아오게 된다. 또한 회전이 멈추어도 내림프가 마찰력에 의해 정지하기까지는 일정시간이 필요하기 때문에 회전이 멈춘 후 cupula는 회전방향으로 편위되어 신경신호를 형성하게 된다[3].
이와 같은 현상은 회전의자를 통한 계단속도검사(stepvelocity rotation)와 같은 지속적인 등속도 회전자극에서 회전 중에 전정신경의 발화(neural firing)와 안진이 줄어들어 없어지고 회전 직후에는 반대방향으로 발화와 안진이 발생하는 현상으로 확인할 수 있다[15,16]. 이 현상은 매우 짧은, 예를 들면 1–2초 내의 회전에서도 발생한다[1]. 따라서 뇌에 전달되는 속도 자극은 부정확하게 된다(Fig. 3).
2. 선가속도 측정에 있어 이석 기능의 불완전성
이석기관은 선가속도계(linear accelerometer)로 중력가속도(gravitational acceleration)를 측정하고 이동에 동반된 관성가속도(inertial acceleration)를 측정한다. 관성가속도는 이동가속도(translational acceleration)와 크기가 같고 방향이 반대이다. 중력가속도는 변하지 않는 크기의 가속도이며 관성가속도는 시시각각 변한다. 우리는 지구 위에서 바로 서 있다는 인식을 유지하기 위해 중력가속도의 방향에 대한 정확한 정보가 필요하다. 또한 선형 이동에 따른 인식, 안반사, 체간 및 자율신경반사를 만들어 내기 위해서는 이동에 따른 관성가속도를 측정해야 한다. 그러나 생리적 선가속도계인 이석기관은 등가 원리의 법칙(equivalent principle; 어떤 선가속도계도 중력가속도와 관성가속도를 구분하지 못한다)에서 제시된 바와 같이 중력가속도와 관성가속도를 구분해내지 못하고 두 가지의 벡터합인 중력관성가속도(gravitoinertial acceleration, GIA)를 측정해 뇌에 전달한다[6,17]. 이를 GIA의 모호함(ambiguity of GIA) 또는 기울임과 이동의 모호함(ambiguity of tilt and translation)이라고 한다(Fig. 4).
중추전정신경계의 기능(VELOCITY-STORAGE CIRCUIT)
말초전정신경계에서 전달되는 신경신호의 부정확성과 모호성에도 불구하고, 우리는 일상 생활에서 정확한 회전과 이동에 대한 인식을 가지고 중력에 대한 위치 인식을 유지한다. 이는 중추전정신경계가 말초전정신경계의 약점을 보완함을 암시한다. 이를 쉽게 이해하기 위해 먼저 내적 모델(internal model)에 대한 내용을 잠시 알아보고 중추신경계의 작동원리를 설명하겠다.
1. 내적 모델(Internal Model)
제어이론에서 내적 모델이란 시스템 변동의 결과를 예측하기 위해 시스템의 반응을 시뮬레이션(simulation)하는 과정을 의미한다. 내적 모델은 시스템이 장치(plant)와 조절자(controller)의 지속적인 상호작용에 의해 조절된다고 가정하며, 인간의 운동 시스템에 대입하면 장치는 몸의 일부가 될 것이고, 조절자는 신경 시스템이 된다. 인간의 운동 시스템에서 내적 모델은 운동명령에 의한 몸의 반응을 예측하고 더 나은 운동반응을 이끌어내는 데 작용한다. 내적 모델은 운동명령에만 국한되지 않고 감각계에도 적용할 수 있다[18]. 내적 모델은 신경 시스템에서보다 구체적인 의미를 지니는데, 신경 발화가 운동명령과 물리적 자극에 대한 감각, 더 나아가 물리적 법칙(예를 들면 f=ma, dg/dt=−Ω×g)도 대변하는 것으로 본다. 이와 같은 개념은 신경반응을 예측하는데 우리가 일상적으로 사용하는 물리적 법칙을 적용할 수 있음을 의미한다[19]. 내적모델에 기반하여 중추전정신경계가 물리적 법칙을 수행한다는 방식을 설명하고 이에 대한 실험적 근거를 소개하고자 한다.
2. 회전 속도의 측정(HV=CV+k×∫CV dt)
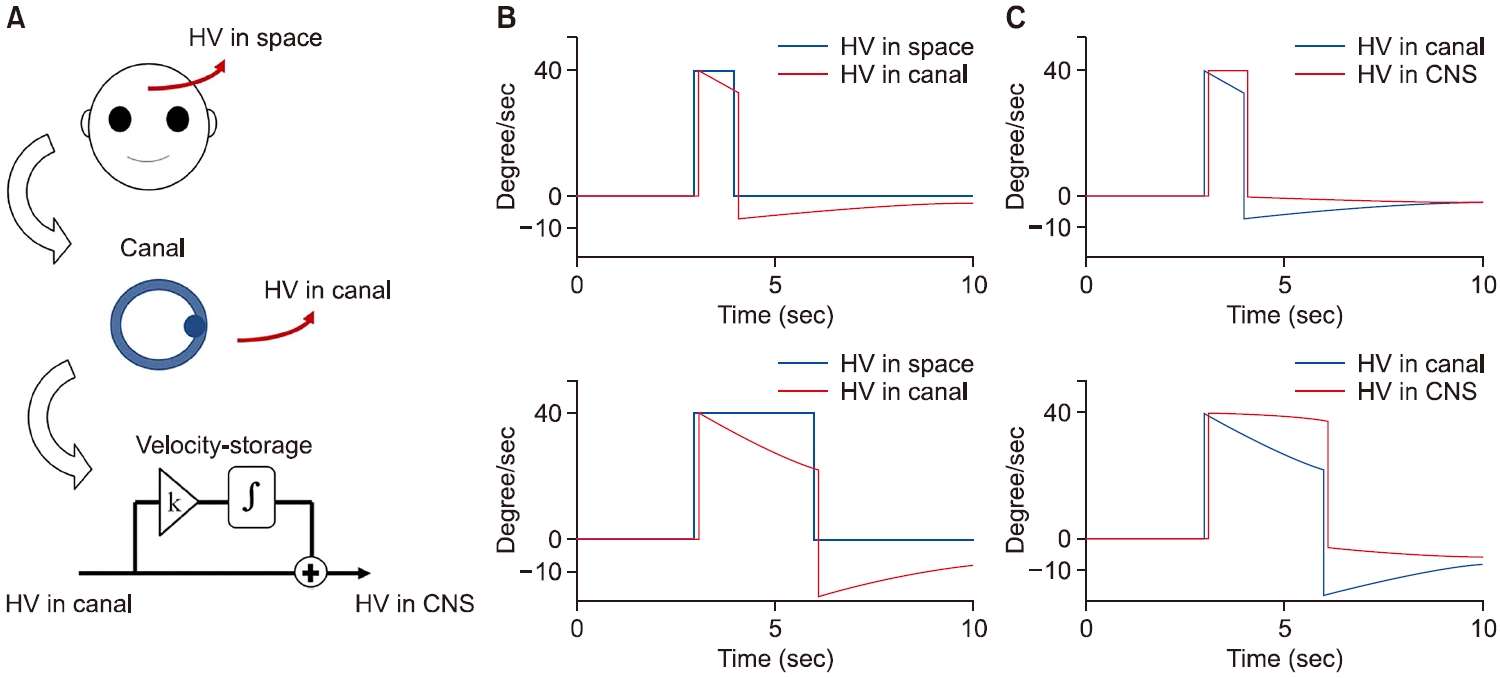
반고리관은 회전 중 그리고 회전 직후 신경신호의 부정확성을 가지고 있다. 그러나 중추전정신경계는 반고리관 신호를 이용하여 정확한 회전속도를 형성한다. 이제 그 원리를 내적 모델에 근거하여 살펴보겠다. 머리가 공간에서 회전하는 속도를 head velocity (HV), 내림프가 공간에서 회전하는 속도를 endolymph velocity (EV), 그리고 반고리관의 신경신호를 canal velocity (CV)라고 하자. CV는 머리의 회전속도와 내림프의 회전속도의 상대적 차이에 의해 cupula가 편위되면서 만들어진다. 이를 식으로 표현하면 CV=HV–EV (Eq. 1)이다. 우리가 알고자 하는 것은 HV이므로, HV=CV+EV (Eq. 2)로 기술하면 된다. CV는 뇌로 전달되어 알고 있으므로 EV만 알면 된다. EV는 내림프의 가속도(endolymph acceleration, EA)를 적분하여, 즉 EV=∫(EA) dt (Eq. 3)을 통해 구할 수 있다. 내림프와 막성미로와의 마찰력(friction)에 의해 내림프가 이동하기 때문에 내림프의 가속도는 마찰력에 비례한다. 즉 EA∝frictional force이며 식을 세우면 EA=k1×frictional force (Eq. 4)이다. 그리고 이동마찰력은 속도 차이에 비례하기 때문에 (frictional force∝HV–EV), 이를 식으로 구성하면 Frictional force=k2×(HV–EV)=k2×(CV) (Eq. 5)이다. 따라서 Eq. 4와 Eq.5를 합치면, EA=k1×k2×(CV)가 되며 k1×k2를 k3로 치환하면 EA=k3×(CV)가 된다. 이제 이를 Eq. 3에 대입하면 EV=∫k3×(CV) dt=k3×∫CV dt가 되며 이를 다시 Eq. 2에 대입하면 HV=CV+k3×∫CV dt가 된다[1]. 즉 우리 중추전정신경계는 (1) 반고리관 정보와 (2) 그 정보의 크기를 조정하여 적분한 값을 더하여 공간에서 실제 이동한 회전속도를 추정할 수 있는 것이다(Fig. 3).
그렇다면 적분이라는 수학적 과정을 신경신호가 만들어낼 수 있어야 한다. 해부학적으로 전정신경핵은 1형과 2형 신경세포를 가지며, 이들은 교차섬유를 통해 반대측 전정신경세포와 연결된다[3,16]. 각 신경세포의 연결과정과 연결세포에 미치는 영향을 시간적으로 따라가보면, 일측 전정신경핵에 전달된 속도신호가 시간차를 두고 반복적으로 되먹임을 하고 있다는 것을 알 수 있다(Fig. 5). 속도저장회로라고 부르는 이 회로는 수학적 적분과정을 수행하고 있는 것을 알 수 있다. 마지막으로 k3라고 말한 상수는 개인마다, 그리고 질병 상태에 따라 다르게 설정될 수 있을 것이다.
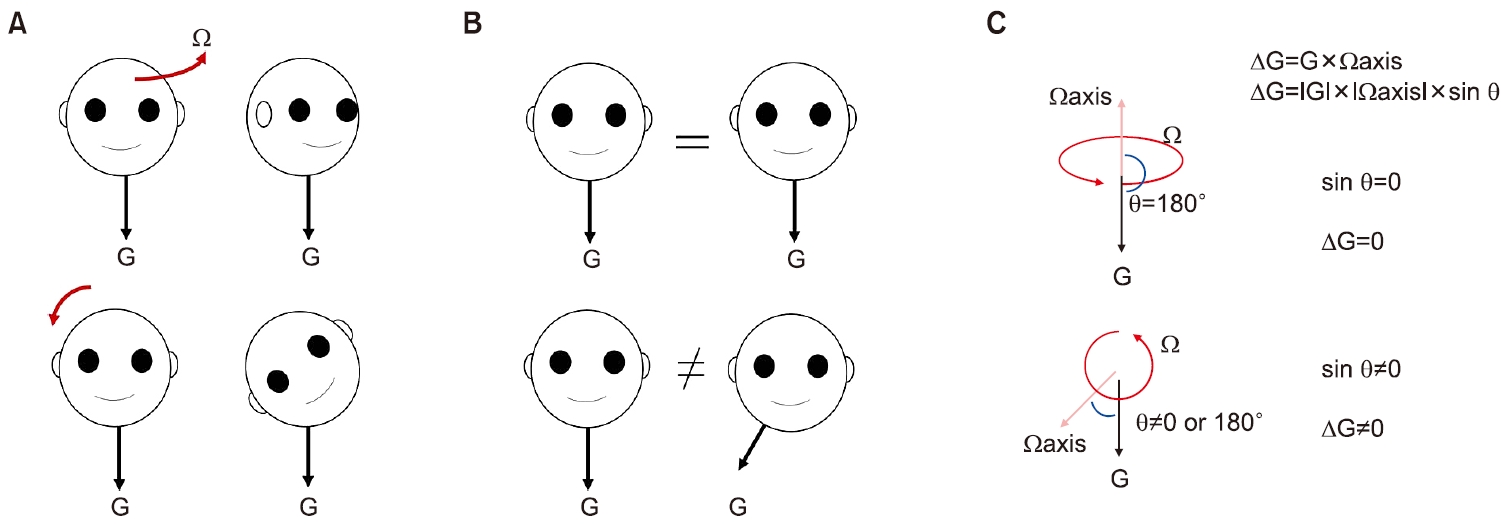
3. 중력가속도의 측정(∫g×Ω dt)과 관성가속도의 계산(inertia=GIA–g)
이석기관은 중력가속도와 관성가속도의 벡터합인 GIA를 뇌에 전달한다. 그렇다면 우리 뇌는 일상적인 움직임 중에서 어떻게 중력가속도에 대한 방향을 정확하게 인식할 수 있을까? 두 가지 특성을 기억할 필요가 있다. 하나는 중력가속도의 크기는 일정하다. 다른 하나는 머리를 숙이거나 기울여 머리중심좌표계(head centered reference frame)에서 중력의 방향이 변하면 반드시 회전이 발생한다는 점이다. 우리는 앞서 중추전정신경계가 어떻게 정확한 회전속도를 측정하는지 살펴보았다. 그리고 회전속도는 벡터량으로 표시할 수 있다. 중력의 방향은 중력의 변화량을 시간에 따라 적분하면 계산할 수 있는데 이를 식으로 작성하면 g=∫(dg/dt) dt (Eq. 6)이다. dg/dt는 시간에 따른 중력벡터의 변화량(중력의 시간에 대한 미분값)으로 회전 벡터(Ω)와의 외적(cross product)으로 표시할 수 있다(dg/dt=g×Ω; Eq. 7). Eq. 6와 Eq. 7을 합치면 g=∫g×Ωdt로 쓰게 된다[1]. 회전 벡터 Ω와 중력 벡터 g가 일직선에 정렬하면 사이각이 0° 또는 180°가 되고, 외적 공식(|g|ㆍ|Ω|ㆍsinΘ)에 의해 dt/dg=0이고 변화량이 없으니 g의 방향도 변화가 없다. 다시 말해서 우리가 바로 서있는 자세에서 수평회전을 할 때 머리중심좌표계에서 중력의 방향은 변하지 않는다. 반면 옆으로 기울이게 되면 사이각이 0° 또는 180° 이외의 값으로 형성되어 dt/dg≠0이 되고, 이를 적분하여 새로운 중력방향을 설정할 수 있다. 예를 들면 머리를 공간에서 오른쪽 어깨 쪽으로 기울이면 머리중심중심좌표계에서는 중력의 방향이 우측 귀 쪽으로 이동하는 것을 알 수 있다(Fig. 6). 이와 같은 방식으로 중력가속도의 방향을 측정하게 되면, 그 이후 관성가속도는 이석기관에서 전해진 GIA에서 중력가속도를 감하면 구할 수 있다(inertia=GIA–g) [6].
4. 신경연산의 증거
신경연산이 실제로 이루어지는지, 그렇다면 어디서 그 역할을 담당하는지에 대한 많은 연구가 진행되고 있다. 가장 대표적인 원숭이를 통한 실험연구 한가지를 소개하고자 한다[20-22]. 앞서 머리를 좌측 어깨 쪽으로 roll 회전시키는 머리기울임(tilt)과 우측으로의 선형이동(translation)은 이석기관에서 동일한 신경신호를 발생시킬 수 있다고 하였다. 그 크기가 같게 머리기울임과 선형이동자극을 맞추고, (1) 선형이동자극 단독, (2) 기울임자극 단독, (3) 기울임자극–선형이동자극, (4) 기울임자극+선형이동자극을 설정하였다. (1)은 이동가속도에 의한 이석기관의 신호(a)만 파생하며, (2)는 기울임에 의한 이석기관의 신호(a), 그리고 반고리관 신호(Ω)가 같이 발생하며, (3)은 이석기관의 신호는 상쇄되고 반고리관 신호(Ω)만 존재하고, (4)의 경우 이동가속도와 기울임에 의한 신호가 합쳐져 2배가 되며(2×a) 반고리관 신호가 존재한다. 이와 같은 자극을 정상인 상태와 반고리관을 막아 회전신호를 소실시킨 상태에서 주고 결절과 목젖의 푸르키네(Purkinje) 세포의 simple spike활성을 측정한 연구가 있다.
연구 결과는 정상에서 결절과 목젖의 푸르키네 세포의 simple spike는 (1), (3), (4)에서 동일하고 (2)에서는 활성이 없었다. 즉 중력방향이 변하는 기울임자극에 대해 푸르키네 세포의 simple spike가 반응하지 않는 반면 이동에 의한 관성가속도에 대해서는 충실히 반응한다는 것을 보여주었다. 또한 반고리관 회전신호를 소실시킨 상태에서는 (1)과 (2)는 동일한 활성을, (3)에서는 반응이 없고, (4)에서는 활성이 2배가 됨을 관찰하였다. 즉 회전신호가 없는 상태에서는 Ω의 벡터가 0이 되고 그에 따른 중력 변화량도 0이 되어 이석기관에서 전달되는 GIA 신호의 변화량을 모두 관성가속도로 측정함을 시사한다. 이를 통해 앞서 설명한 inertia=GIA–g의 내적 모델이 입증되는 것을 알 수 있고, 결절과 목젖에서 관성가속도를 만들어내는 것을 알 수 있다. 또한 (3)의 반응, 즉 이석신호는 없고 반고리관에서 roll 회전신호만 존재하는 신경반응과 바로 서거나 누워서 중력가속도 방향에 정렬된 회전자극 시 나타나는 신경반응을 비교하여 결절과 목젖이 중력가속도 방향에서 정렬되지 않은 회전만 반영하는 것을 확인하였다. 그리고 이 반응의 크기와 시간지연을 자극 속도와 비교하여 신경반응이 회전속도가 아닌 회전위치를 나타냄을 증명하였다. 이는 소뇌의 결절과 목젖이 반고리관 신호를 받아 머리기울임(중력방향)을 추정 후 이석기관에서 전달된 GIA에서 중력가속도를 감하여 관성가속도를 표현함을 시사하는 소견이 된다[20-22].
5. 속도저장회로 내의 2개의 보완적 Feedback Loop
앞서 기술한 속도저장회로에서도 두 가지 문제점이 있다. 한 가지는 어떻게 중력에 대한 초기 인식을 가지게 되었을까 하는 것이다. 처음 중력을 느꼈을 때, 예를 들면 우주비행 후 지구에 돌아왔을 때, 우리 뇌는 중력이 없는 환경에서 중력이 있는 환경으로 돌아오는데, 중력과 관성을 구분할 수 없기 때문에 중력을 인식하기 위한 방법이 필요할 수 있다. 이를 설명하는 것이 somatogravic feedback의 개념이다. 즉 지속적이고 변하지 않는 관성을 중력으로 인식하는 것이다. 이와 같은 현상을 설명하는 것이 somatogravic illusion인데, 예를 들어 등가속운동을 하는 물체에서 우리는 종종 기울었다는 느낌을 받게 된다[23]. 중심축에서 벗어나 지속적으로 회전(off-axis vertical rotation)할 때 중심을 향한 구심력이 있으며, 이석기관은 원심력 방향으로 이를 측정한다. 이때 발생하는 기울임인식(tilt perception)도 같은 현상이다[24,25].
다른 하나의 문제점은 반고리관을 통해 측정된 회전신호의 불완전성이다. 예를 들어 반고리관의 손상이 발생하게 되면, 이를 통해 계산된 중추전정신경계의 회전속도도 부정확하게 된다. 정상에서도 일정기간의 회전 후에는 회전 후 반응이 발생한다. 이 또한 중력 추정에 문제를 야기하게 된다. 이 문제를 극복하기 위해 중추신경계에서는 추가적 회전속도를 파생하여 중력가속도의 위치를 GIA의 방향에 일치시키는데, 이를 rotational feedback이라고 한다. 이에 대한 증거를 바비큐 회전자극을 통해 알아볼 수 있다. 옆으로 누워서 한 방향으로 계속 회전을 가정하자. 이동이 없으므로 이석에서 전달된 GIA는 중력과 같다. 처음에는 반고리관이 회전자극을 충실히 전달하기 때문에 이석에서 전달된 GIA와 뇌에서 추정된 중력의 방향이 같게 된다. 그러나 1분 이상 회전하게 되면 반고리관은 더이상 회전신호를 파생하지 못하게 되고, 따라서 중력가속도의 방향을 추정하는데 문제가 생기게 된다. 따라서 중력 위치에 대한 인식에 오류가 생기고 이석기관에서 전달된 GIA 벡터와 뇌에서 추정한 중력가속도 벡터가 일치하지 않게 된다. 이는 가성관성가속도를 우리 뇌가 계산하게 한다. 그러나 중추신경계는 rotational feedback을 통해 중력가속도를 GIA에 맞추고자 하기 때문에 지속적인 회전자극을 만들어 중력에 대한 올바른 인식을 유지한다. 실제 바로 선 자세에서 회전하는 것과 다르게 바비큐 회전에서는 안진이 지속적으로 관찰된다. 수직축 이탈회전(off-vertical axis rotation)에서 관찰되는 안진의 bias가 이와 같은 현상이다[26,27].
6. 속도저장회로 요약
정리하면, 중추전정신경계는 양측 전정신경핵과 이를 연결하는 교차섬유, 소뇌의 결절과 목젖으로 구성된 속도저장회로를 통해 말초전정신경에서 전달된 부정확한 회전속도 신호와 모호한 GIA를 정제하고 구분하는 역할을 한다. 이렇게 발생된 신경신호는 앞서 언급한 전정인식, 전정안반사, 전정척수반사, 그리고 전정자율신경반사에 사용되게 된다.
전정질환에서의 임상증상 및 징후를 속도저장회로의 측면에서 해석
앞서 우리는 전정신경계의 해부학적 구조와 목적에 대해 알아보았다. 이제 속도저장회로의 생리적 기능을 이해함으로써, 전정신경계 질환자에서 보이는 임상증상 및 징후를 어떻게 해석할 수 있는지 알아보겠다. 대표적으로 인식과 눈운동, 그리고 자율신경반사의 측면으로 소개하고자 한다.
1. 양성돌발체위현기증에서 어지럼의 특징: 선형가속도 인식의 증거
양성돌발체위현기증(benign paroxysmal positional vertigo)은 이석기관에서 유리된 이석 부스러기가 반고리관에서 흘러 다니며 잘못된 회전속도를 유발하여 어지럼을 발생시키는 질환이다[28]. 일반적인 양상은 체위에 의해 발생하는 회전성 어지럼이다[29]. 그러나 일부 환자에서는 종종 회전성 어지럼에 동반하여 또는 회전성 어지럼은 없이 떨어지거나 뜨는 느낌의 선형 어지럼(linear vertigo)을 호소하는 경우가 있다. 실제 환자의 증상을 전향적으로 수집한 연구에서 이와 같은 체위 변화 후 발생한 선형 어지럼은 약 35%에서 관찰되었다[30]. 해당 연구에서는 체위 변화 후 발생한 가성 회전자극(false rotational cue)에 의해 중력가속도의 측정에 오류가 발생하고 결과적으로 잘못된 선형가속도를 중추신경계가 만들어내어 이를 인식하게 된다고 가정하였다. 속도저장 기전을 구현한 모델을 통해 양성돌발체위현기증에서 발생하는 가성 관성가속도를 구현하고 그 방향이 실제 환자가 호소하는 선형 어지럼의 방향과 일치하는 것을 보여주어(Fig. 7), 환자의 선형 어지럼을 설명할 수 있었다[30].
2. 중추성 체위현훈의 안진 특성
중추성 체위현훈은 뇌간 및 소뇌의 뇌병변으로 인해 체위 변화 시 발생하는 어지럼과 안진을 특징으로 한다. 어지럼이 강하고 안진이 짧게 관찰되는 경우에 중추성 돌발체위현훈(central paroxysmal positional vertigo)이라고 구분하며 어지럼은 심하지 않으나 지속적인 안진이 관찰되는 경우 중추성 지속체위안진(central persistent positional nystagmus)이라고 한다[31].
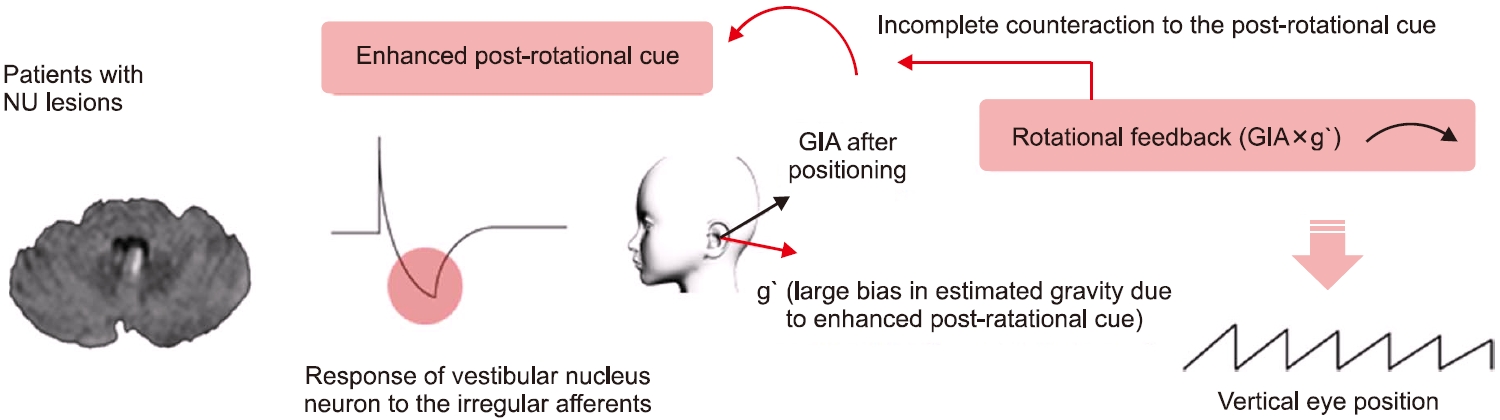
중추성 돌발체위현훈을 분석한 연구에서 안진의 방향이 머리의 회전 후 즉각적으로 발생하며, 머리 회전의 반대방향을 향하고 약 5초의 시간 상수를 가지면서 감쇠하는 특징을 확인하여 반고리관 신호에서 근원하는 안진임을 제시하였다[32,33]. 앞서 속도저장회로의 보완적 feedback loop 중 rotational feedback은 이러한 안진을 억제하는 기능이 있다. 그러나 중추신경계, 특히 소뇌에 병변이 있는 경우, 전정신경핵의 활성이 증가하여 결과적으로 회전 후 반응이 증가되며, rotational feedback의 기능도 원활히 적절히 작용하지 못하게 되므로 회전 후 안진이 중추성 돌발체위현훈에 동반하여 발생하는 것을 제시한 바 있다(Fig. 8).
중추성 지속체위안진은 특정 자세에서 시간에 따라 감쇠하지 않는 지속성을 보이는데, 중력의 방향에 따라 그 크기가 변화하는 특징도 있다. 최근 연구에서는 소뇌 병변에 의해 중력방향 추정에 지연 및 오류(bias)가 발생할 것을 가정하였고(Fig. 9), 이는 실제 이석기관에서 전달되는 GIA와 추정된 중력과의 차이를 유발하여 rotational feedback이 작동되고, 결과적으로 수직축 이탈회전에서 관찰되는 안진의 bias와 같이 지속체위안진을 형성하는 것으로 제시하였다[34].
3. 전정실신
앞서 전정자율신경반사는 이석기관과 교감신경계의 반응이 주요하고, 압력반사와는 다르게 100–200 msec의 짧은 반감기를 가짐을 기술한 바 있다[12-14]. 일반적으로 기립자극 시 체액이 체내의 아래쪽으로 몰리게 된다. 기립자극이 발생하면 실제 머리의 위치는 위로 이동하기 때문에 이석기관은 중력가속도와 아래로 향하는 관성가속도의 합을 뇌에 전달한다[1,17]. 뇌는 중력가속도의 크기를 알고 있으므로, 관성가속도만을 계산할 수 있고, 이 관성가속도는 feedforward 방식으로 교감신경계를 흥분시켜 혈관을 수축시키고 심박수와 심박 출력을 증가시켜 혈압을 일차적으로 보상한다. 만약 그 반응이 부족하게 되면, 압력수용체가 부교감 활성을 떨어트려 혈압을 유지하게 될 것이다. 그러나 만약 이석기관에서 잘못된 관성가속도신호가 발생한다면 잘못된 교감신경 반응을 초래하여 체액의 상대적 변화 없이 혈관 수축 및 심박수와 심박출력 증가를 일으킬 것이고, 이는 비정상적 혈압 상승을 유발할 수 있다. 이때 뒤를 잇는 압력반사는 부교감 활성을 증가시켜 혈압을 떨어뜨리려 할 것이며, 만약 그 반응이 강하다면 미주신경성 실신을 유발할 수 있을 것이다[35]. 또한 속도저장회로의 기능을 고려하면 잘못된 관성가속도는 반드시 이석기관에서 기인하는 것이 아닌 중력가속도에 정렬하지 않는 회전 벡터를 초래하는 반고리관 또는 중추신경계 질환에서도 관찰될 수 있다(Fig. 10).
결 론
전정신경계의 목적과 중추신경 네트워크, 말초전정신경계의 특성과 이를 보상하는 속도저장 기전의 생리적 이해는 전정안반사를 넘어 전정인식, 전정(목)척수반사, 전정자율신경반사에 걸친 이해를 증진시키고 전정질환자의 다양한 임상증상을 해석하는 데 도움이 될 것이다.
ARTICLE INFORMATION
-
저자는 이 논문과 관련하여 이해관계의 충돌이 없음을 명시합니다.
Acknowledgments
본 연구는 한국연구재단의 2020년도 이공학 개인기초연구지원사업의 지원에 의하여 수행되었습니다(과제고유번호 2020R1A2C4002281).
Fig. 1.Head stabilization mechanism during motion. (A) The directions of vestibulo-collic reflex (VCR) and cervicocollic reflex (CCR) during whole body rotation. (B) The proposed model for head stabilization behavior in space and on body. VSR, vestibulospinal reflex.

Fig. 2.Schematic diagram illustrating vestibulo-sympathetic reflex that participates cardiovascular homeostasis. BP, blood pressure.
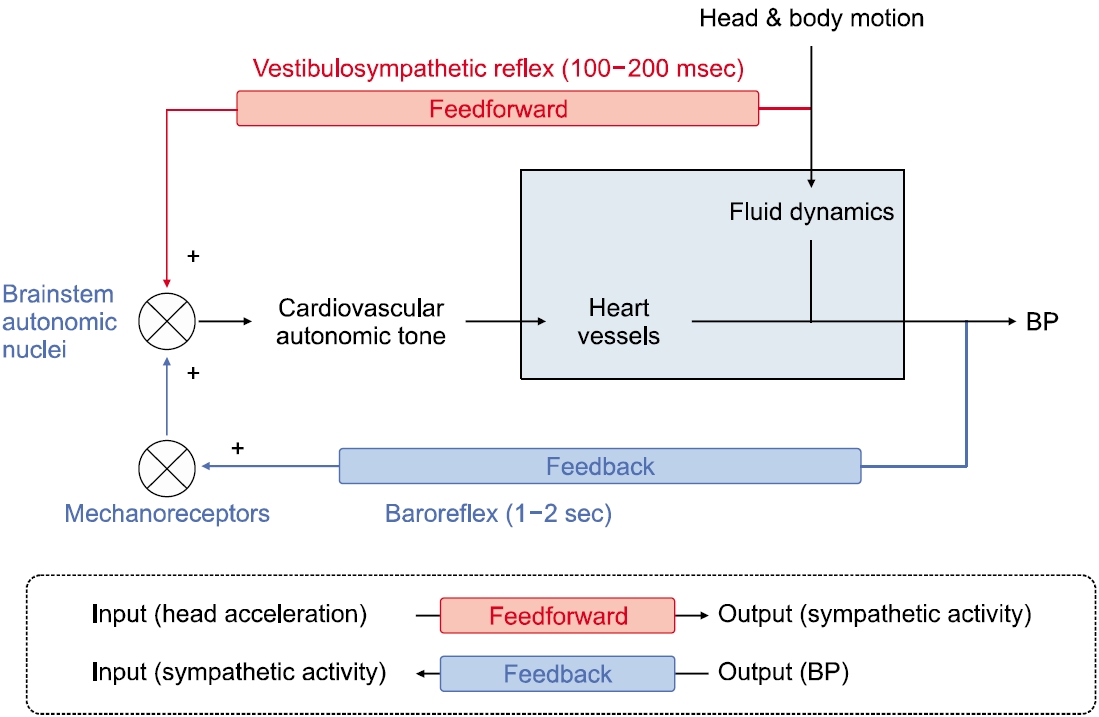
Fig. 3.Rotational head velocity (HV) estimation. (A) The process for rotational velocity estimation. (B) The difference between HV in space and HV in canal during 40°/sec rotation with durations of 1 second (upper) and 3 seconds (lower). (C) Improvement of rotational velocity estimation in central nervous system (CNS).
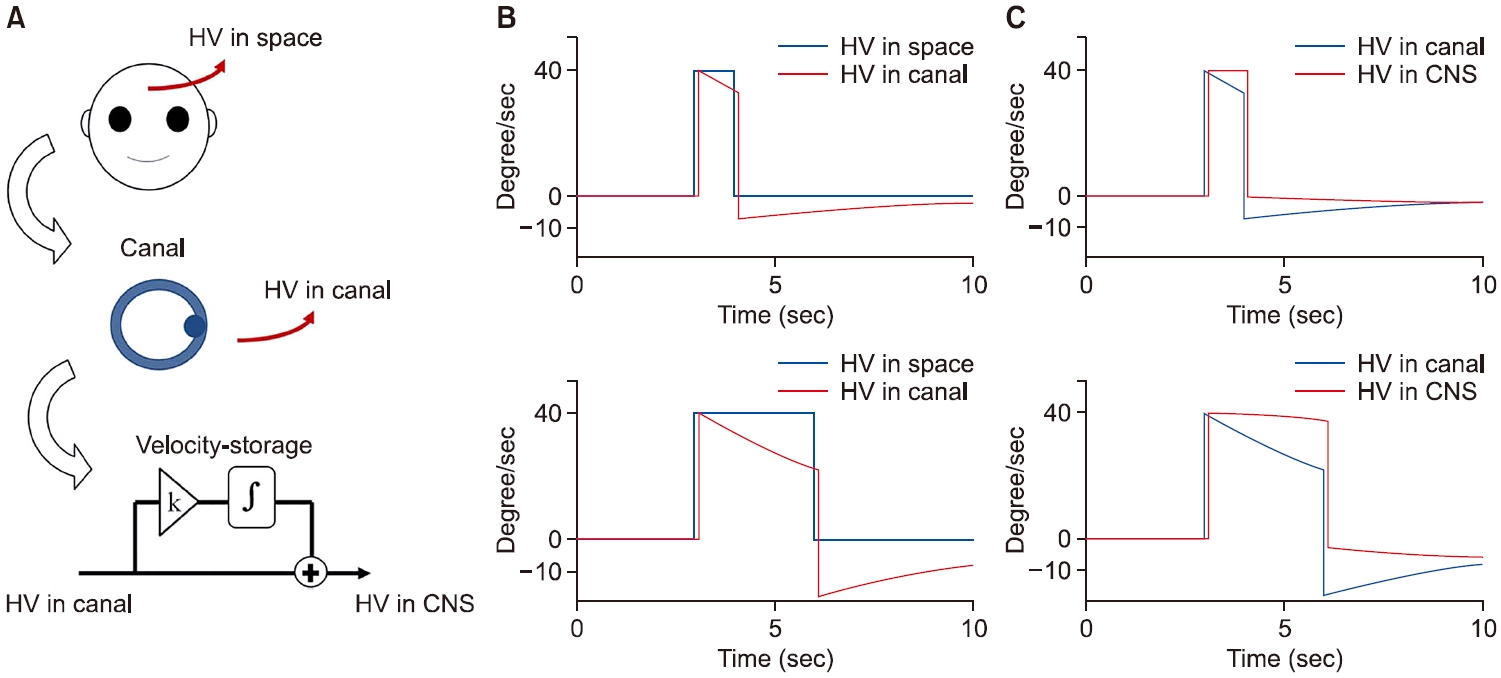
Fig. 4.Ambiguity of tilt and translation. Ω, rotational velocity; GIA, gravitoinertial acceleration; G, gravitational acceleration.

Fig. 5.Integration of velocity signals over time within velocity-storage circuit.
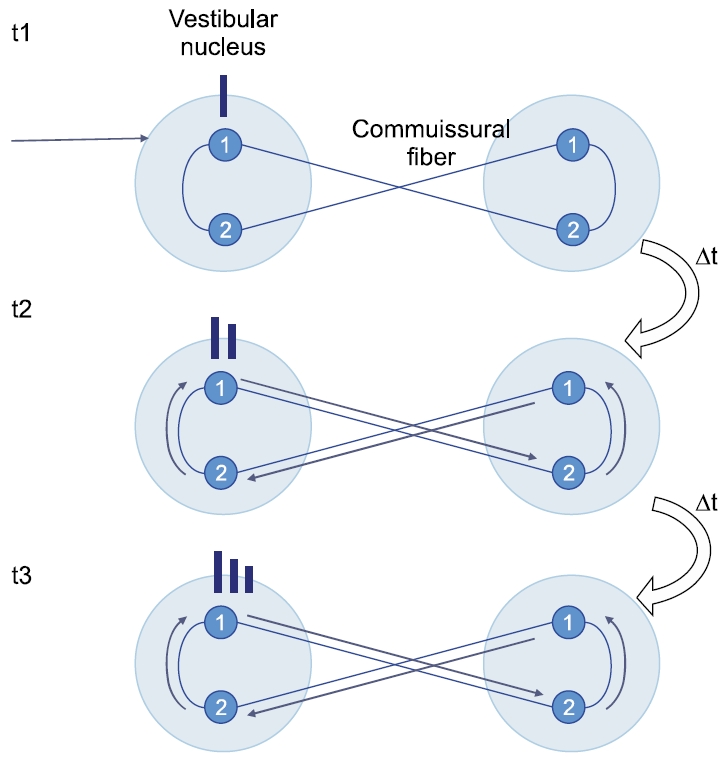
Fig. 6.Gravity estimation by using rotational and otolith cue. (A) Yaw (upper) and roll (lower) rotations in earth reference frame. (B) Internal representation of gravity. (C) Internal model estimating the change of gravity direction. G, gravity.
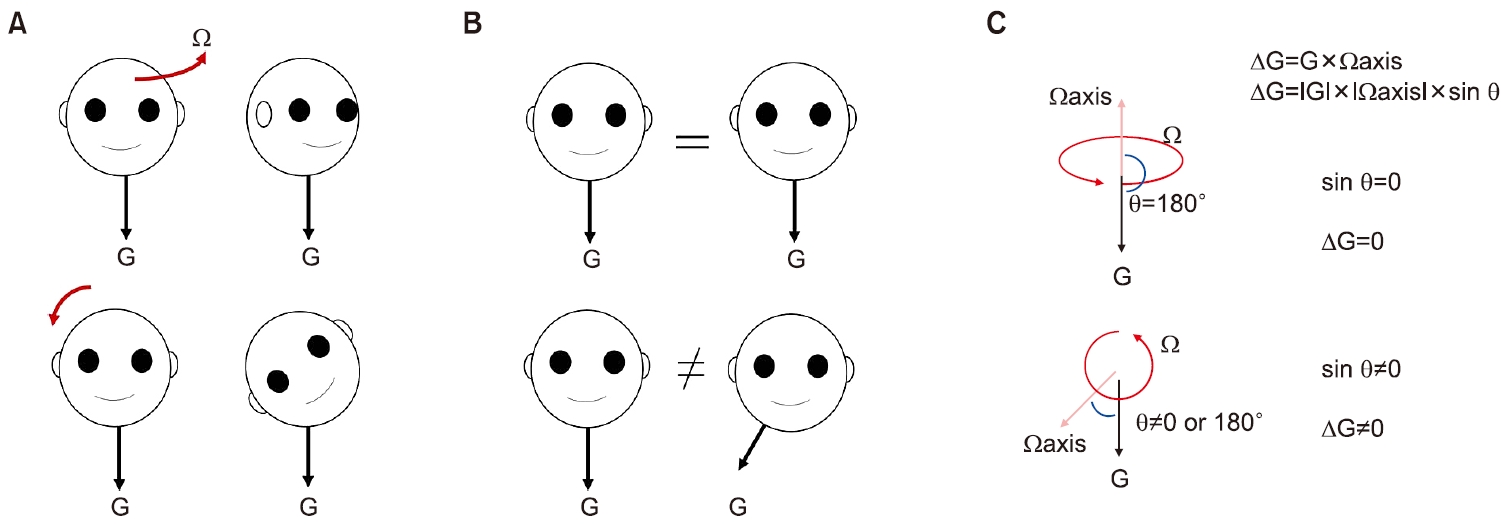
Fig. 7.The prevalence and the mechanism of liner vertigo in patients with benign paroxysmal positional vertigo (BPPV). (A) The prevalence of linear vertigo in patients with posterior and horizontal canal BPPV. The linear vertigo was reported in 34.4% of patients with BPPV (upper). In terms of severity (lower), the patients reported to have a fear from linear vertigo in 28.6%. (B) The direction of linear vertigo in patients with posterior canal BPPV (upper) and horizontal canal BPPV (lower). (C) The simulated direction of inertia induced by false RC (upper, posterior canal BPPV; lower, horizontal canal BPPV) was concordant with the direction of linear vertigo reported from the patients. RV, rotational vertigo; LV, linear vertigo; RC, rotational cue.
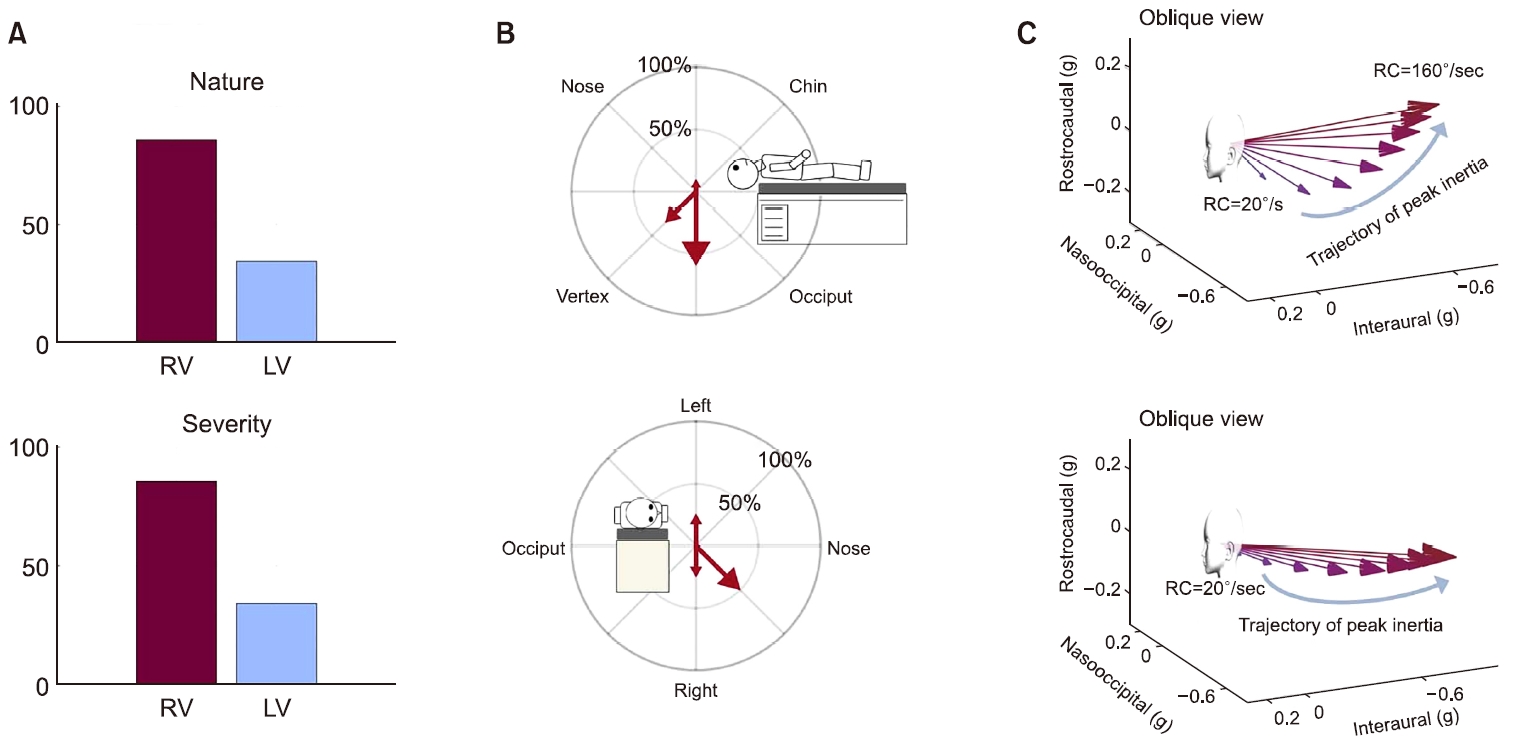
Fig. 8.The mechanism of central paroxysmal positional nystagmus. NU, nodulus and uvula; GIA, gravitoinertial acceleration.
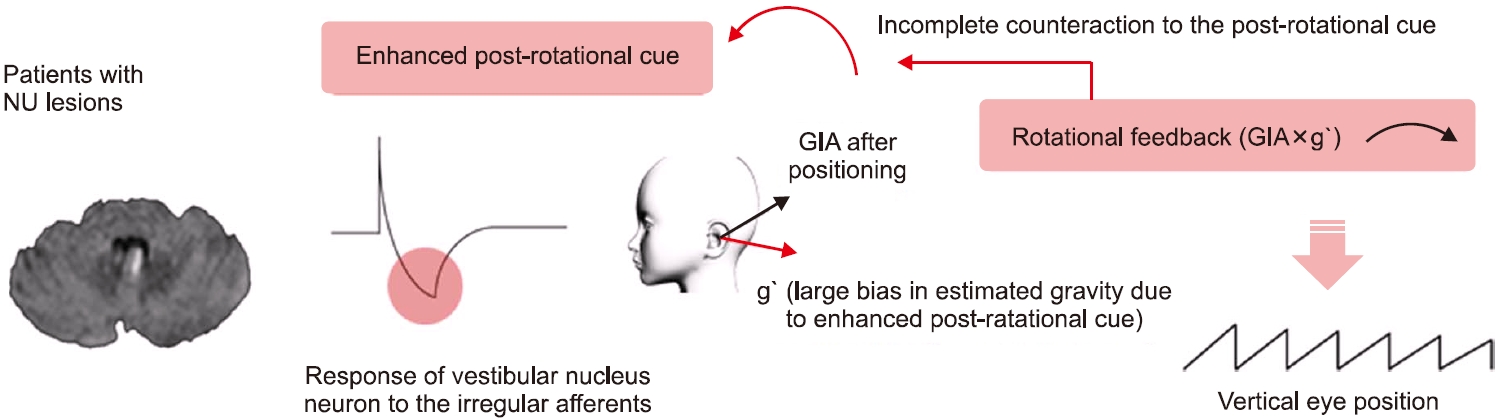
Fig. 9.The mechanism of central persistent positional nystagmus. (A) Schematic diagram representing the lesion induced gravity estimation bias within the velocity-storage circuit. (B) Model simulation showed persistent false rotational cue (estimated w) induced by central lesions without rotational cue during ear-down positions. GIA, gravitoinertial acceleration; CPN, central positional nystagmus.
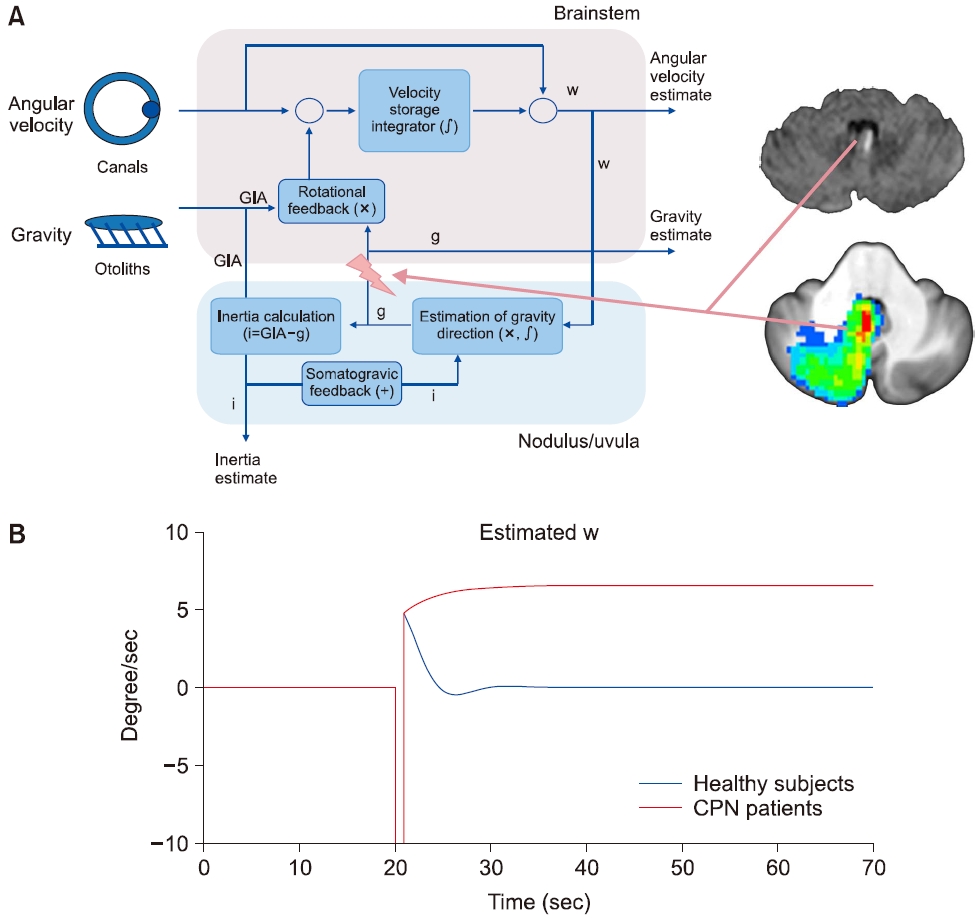
Fig. 10.The autonomic response during orthostatic maneuver and the expected response in vestibular syncope. GIA, gravitoinertial acceleration; BP, blood pressure.
REFERENCES
- 1. Laurens J, Angelaki DE. The functional significance of velocity storage and its dependence on gravity. Exp Brain Res 2011;210:407–22.ArticlePubMedPMC
- 2. Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodal sense. Annu Rev Neurosci 2008;31:125–50.ArticlePubMed
- 3. Leigh RJ, Zee DS. The neurology of eye movements 5th ed. New York, Oxford University Press. 2015.
- 4. Seemungal BM. The cognitive neurology of the vestibular system. Curr Opin Neurol 2014;27:125–32.ArticlePubMed
- 5. MacNeilage PR, Banks MS, DeAngelis GC, Angelaki DE. Vestibular heading discrimination and sensitivity to linear acceleration in head and world coordinates. J Neurosci 2010;30:9084–94.ArticlePubMedPMC
- 6. Wexler M, Panerai F, Lamouret I, Droulez J. Self-motion and the perception of stationary objects. Nature 2001;409:85–8.ArticlePubMed
- 7. Dupin L, Wexler M. Motion perception by a moving observer in a three-dimensional environment. J Vis 2013;13:15. Article
- 8. Beylergil SB, Shaikh AG. Visual perception of heading in the syndrome of oculopalatal tremor. Cerebellum 2020 Aug 1 [Epub]. https://doi.org/10.1007/s12311-020-01176-w.Article
- 9. Goldberg JM, Cullen KE. Vestibular control of the head: possible functions of the vestibulocollic reflex. Exp Brain Res 2011;210:331–45.ArticlePubMedPMC
- 10. Peterson BW, Goldberg J, Bilotto G, Fuller JH. Cervicocollic reflex: its dynamic properties and interaction with vestibular reflexes. J Neurophysiol 1985;54:90–109.ArticlePubMed
- 11. Peng GC, Hain TC, Peterson BW. Predicting vestibular, proprioceptive, and biomechanical control strategies in normal and pathological head movements. IEEE Trans Biomed Eng 1999;46:1269–80.ArticlePubMed
- 12. Jian BJ, Acernese AW, Lorenzo J, Card JP, Yates BJ. Afferent pathways to the region of the vestibular nuclei that participates in cardiovascular and respiratory control. Brain Res 2005;1044:241–50.ArticlePubMed
- 13. Yates BJ, Bronstein AM. The effects of vestibular system lesions on autonomic regulation: observations, mechanisms, and clinical implications. J Vestib Res 2005;15:119–29.ArticlePubMed
- 14. Raphan T, Cohen B, Xiang Y, Yakushin SB. A model of blood pressure, heart rate, and vaso-vagal responses produced by vestibulo-sympathetic activation. Front Neurosci 2016;10:96. ArticlePubMedPMC
- 15. Waespe W, Henn V. Neuronal activity in the vestibular nuclei of the alert monkey during vestibular and optokinetic stimulation. Exp Brain Res 1977;27:523–38.ArticlePubMed
- 16. Wearne S, Raphan T, Cohen B. Contribution of vestibular commissural pathways to spatial orientation of the angular vestibuloocular reflex. J Neurophysiol 1997;78:1193–7.ArticlePubMed
- 17. Merfeld DM, Zupan L, Peterka RJ. Humans use internal models to estimate gravity and linear acceleration. Nature 1999;398:615–8.ArticlePubMed
- 18. Tin C, Poon CS. Internal models in sensorimotor integration: perspectives from adaptive control theory. J Neural Eng 2005;2:S147–63.ArticlePubMedPMC
- 19. MacNeilage PR, Ganesan N, Angelaki DE. Computational approaches to spatial orientation: from transfer functions to dynamic Bayesian inference. J Neurophysiol 2008;100:2981–96.ArticlePubMedPMC
- 20. Yakusheva TA, Shaikh AG, Green AM, Blazquez PM, Dickman JD, Angelaki DE. Purkinje cells in posterior cerebellar vermis encode motion in an inertial reference frame. Neuron 2007;54:973–85.ArticlePubMed
- 21. Angelaki DE, Yakusheva TA. How vestibular neurons solve the tilt/translation ambiguity. Comparison of brainstem, cerebellum, and thalamus. Ann N Y Acad Sci 2009;1164:19–28.PubMedPMC
- 22. Laurens J, Meng H, Angelaki DE. Computation of linear acceleration through an internal model in the macaque cerebellum. Nat Neurosci 2013;16:1701–8.ArticlePubMedPMC
- 23. Paige GD, Seidman SH. Characteristics of the VOR in response to linear acceleration. Ann N Y Acad Sci 1999;871:123–35.ArticlePubMed
- 24. Graybiel A, Johnson WH, Money KE, Malcolm RE, Jennings GL. Oculogravic illusion in response to straight-ahead acceleration of CF-104 aircraft. Aviat Space Environ Med 1979;50:382–6.PubMed
- 25. Merfeld DM, Zupan LH, Gifford CA. Neural processing of gravito-inertial cues in humans. II. Influence of the semicircular canals during eccentric rotation. J Neurophysiol 2001;85:1648–60.ArticlePubMed
- 26. Glasauer S. Interaction of semicircular canals and otoliths in the processing structure of the subjective zenith. Ann N Y Acad Sci 1992;656:847–9.ArticlePubMed
- 27. Angelaki DE, Hess BJ. Three-dimensional organization of otolith-ocular reflexes in rhesus monkeys. II. Inertial detection of angular velocity. J Neurophysiol 1996;75:2425–40.ArticlePubMed
- 28. Kim JS, Zee DS. Clinical practice. Benign paroxysmal positional vertigo. N Engl J Med 2014;370:1138–47.ArticlePubMed
- 29. Furman JM, Cass SP. Benign paroxysmal positional vertigo. N Engl J Med 1999;341:1590–6.ArticlePubMed
- 30. Choi JY, Park YM, Lee SH, Choi J, Hyun SW, Song JM, et al. Linear vertigo in benign paroxysmal positional vertigo: prevalence and mechanism. Cerebellum 2021;20:160–8.ArticlePubMed
- 31. Büttner U, Helmchen C, Brandt T. Diagnostic criteria for central versus peripheral positioning nystagmus and vertigo: a review. Acta Otolaryngol 1999;119:1–5.ArticlePubMed
- 32. Choi JY, Kim JH, Kim HJ, Glasauer S, Kim JS. Central paroxysmal positional nystagmus: characteristics and possible mechanisms. Neurology 2015;84:2238–46.ArticlePubMed
- 33. Choi JY, Kim JS. Nystagmus and central vestibular disorders. Curr Opin Neurol 2017;30:98–106.ArticlePubMed
- 34. Choi JY, Glasauer S, Kim JH, Zee DS, Kim JS. Characteristics and mechanism of apogeotropic central positional nystagmus. Brain 2018;141:762–75.ArticlePubMed
- 35. Kwon E, Lee JY, Kim HJ, Choi JY, Kim JS. Can dyssynergia of vestibulosympathetic and baroreflexes cause vestibular syncope? The hypothesis based on the velocity-storage function. Cerebellum 2021 Jun 22 [Epub]. https://doi.org/10.1007/s12311-021-01296-x.Article
Citations
Citations to this article as recorded by




 PubReader
PubReader ePub Link
ePub Link Cite
Cite